A study of interacting Bayesian recurrent neural
networks with incremental learning
__________________________________________________________________________
En
studie av interagerande Bayesianska artificiella neuronnät med gradvis
inlärning
Christopher Johansson
MSc thesis in computer science,
2D1021
Abstract
This thesis investigates the properties of
systems composed of recurrent neural networks. Systems of networks with
different time-dynamics are of special interest. The idea is to create a system
that posses a long term memory (LTM) and a working memory. The working memory
is implemented as a memory that works in a similar way to the LTM, but with
learning and forgetting at much shorter time scales. The recurrent networks are
used with an incremental, Bayesian learning rule. This learning rule is based
on Hebbian learning. In this thesis there is a thorough investigation of how to
design the connection between two neural networks with different time-dynamics.
Another field of interest is the possibility to compress the memories in the
working memory, without any major loss of functionality. In the end of the
thesis, these results are used to create a system that is aimed at modeling the
cerebral cortex.
Sammanfattning
I denna rapport kommer egenskaperna
hos system uppbyggda av återkopplade neuronnät att undersökas. System
uppbyggda av nätverk som arbetar på skilda tidsskalor är av speciellt intresse.
Målet är att skapa ett system med ett långtidsminne och ett arbetsminne.
Arbetsminnet kommer att realiseras på samma sätt som ett långtidsminne, men det
kommer att arbete med mycket kortare tidsskalor. De återkopplade neurala nätverken kommer att tränas med en
inkrementell, Bayesiansk inlärningsregel. Inlärningsregeln är baserade på
Hebbsk inlärning. I rapporten finns en noggrann undersökning av hur man kan
koppla ihop två neurala nätverk. Jag kommer också att undersöka möjligheten att
komprimera representationen i arbetsminnet, utan att det medför sämre
prestanda. I slutet av rapporten kommer jag att använda resultaten från början
av rapporten, till att skapa ett system
som efterliknar hjärnbarken.
Preface_______________________________
Acknowledgements
and general information
This master project in computer science was
preformed at SANS, Studies of Artificial Neural Systems, a department in NADA,
Numerical Analysis and Computer Science, at the Royal Institute of Technology,
KTH. The work was done during the autumn of year 2000. Professor Anders
Lansner, head of the SANS research group, was the examiner of the project.
I would like to thank Anders Sandberg for his
help and support during the project. Anders S. managed to explain all questions
I had during the work on the project. He also taught me a diverse set of new
knowledge. Anders Lansner woke my interest on the subject of research, and he
has all through the work been encouraging. I also would like thank everyone
else in the SANS group for an inspiring environment; Örjan Ekeberg , Erik
Fransén, Pål Westermark, Peter Raicevic, Anders Fagergren, Jeanette Hellgren
Kotaleski, Alexander Kozolov, Erik Aurell.
1.0 Introduction
It’s fairly clear that the brain
posses several kinds of memory processes. These can be divided into two major
categories, long-term memory (LTM) and short-term memory (STM). This thesis
will focus on how the STM can be constructed. The thesis will also be concerned
with the interaction between LTM and STM and the effects that arise in systems
that are comprised of these two sorts of memories.
Long-term memory processes have
for a long time been considered to reside in the synapses. Long-term
potentiation (LTP) and long-term depression (LTD) have been observed to occur
in synapses. LTP and LDP in the synapses are thought to constitute the
long-term memories that we posses [1]. Based on these observations, parallels between
artificial neural networks and populations of nerve cells have been made. Based
on these ideas, an artificial neural network (ANN) can be used as a memory.
This type of ANN is called attractor networks. Attractor networks have been
suggested to constitute a good model of how the LTM memory works [2-6].
It is a common view in the
research community that short-term storage of memories is based on the current
activity in the brain. In this view each short-term memory resides in the brain
as a mode of an activity-wave [7]. The mode of this activity-wave can swiftly be
changed to accommodate new memories or to forget old memories.
In this thesis, a different view
is adopted of how the short-term memory processes is attained. In the presented
view, the short-term memories are stored in the synapses between neurons. It
means that the short-term memory process is similar to the long-term memory
process, but it work on a shorter time scale. In this thesis, I have used an
attractor network with high plasticity, to simulate the STM.
The main focus of the thesis was
to establish that an attractor network with high plasticity could be used as a
model of STM, and that the STM could function as a working memory.
The most important function of a
STM is to hold information about the current situation. The STM also needs to
be able to swiftly change its associations as new information arrives. The STM
does not need to capture the details of the arriving information. The details
are captured and stored by the LTM.
This means that it’s more important for the STM to make the correct
associations to the pattern in LTM than to be able to store the complete
pattern.
An important question is how the
memories stored in the LTM most effectively can be represented in the STM. The
representation of memories in the STM does not need to be identically to the
representation of the memories in the LTM. A requirement to make this possible
is that there is a distinct connection from each memory in the STM to the
corresponding memory LTM. Several different methods can be used to create the
compressed memories of the STM and associate these STM memories to their
corresponding memories in the LTM.
A central concept is working
memory. Working memory is a concept used in cognitive psychology. The term,
“working memory”, was introduced by Alan Baddely [8]. This thesis shows how the STM, based on the Bayesian
neural network, can be used as a working memory. In systems with a working
memory, short-term variable binding (STVB) can be made. STVB is some times
referred to as role filling. STVB is a basic function, needed to
construct a logical reasoning system.
The basic function of an
auto-associative memory is, as the name suggests, to associate the input to one
of the stored memories. If an input pattern is presented to the auto-associative
memory, the memory will respond with the stored memory that most closely
resembles the input. The associative memories in this thesis were implemented
with attractor networks. Attractor networks are constructed with recurrent
networks of artificial neurons. In the recurrent network, each neuron has
connections to all other neurons. Each
of these connections is equipped with a weight, which controls the influence
between the neurons. The connection weights form a matrix. The attractor network
stores the patterns by altering the values of the weights [9].
The attractor networks were
constructed with artificial neurons. The artificial neural networks were
implemented with a palimpsest,
incremental, Bayesian learning rule [10]. This Bayesian learning rule allows the user to
control the temporal properties of plasticity in the network by the
modification of a single parameter. Sandberg et al at SANS, KTH have developed
the incremental, Bayesian learning rule.
Chapter 2 contains a basic introduction to
cognitive neuroscience. There is a description of how memories are categorized
into explicit and implicit memories. The chapter also contains an overview of
the anatomy of the nervous system. Briefly the anatomy and function of nerve
cells are presented. A short
presentation of artificial neural networks is given. A closer look at attractor
networks and the Hopfield model is made. An overview of associative memories is
given, as they constitute a central concept in this thesis. Then, the
incremental, Bayesian learning rule is presented. A biological interpretation
of Bayesian learning rule is also made. Finally, some interesting concepts that
can be found in auto-associative memory systems are presented.
In chapter 3 the implementation of the Bayesian
network model is presented. The physical realization of the neural networks is
presented along with the choice of parameter values. The basic behavior of
attractor networks based on the Bayesian learning rule is presented. The
environment, in which the networks operate in, is also presented along with how
the networks were tested.
The concern of chapter 4 is to present how a
system can be made out of two networks and how these networks can be made to
cooperate. A few basic design ideas are tried and studied.
The focus of chapter 5 is how the
representation of the memories can be compressed in the STM. A couple of
different alternatives are studied. Then, interest is turned to some important
functions that can be found in a system composed of a LTM and STM.
Finally, in chapter 6, there is a demonstration
of how a working memory can be useful. STM is used as working memory in the
systems. The systems are presented with a task that requires the use of both a
LTM and STM. It is also shown how larger systems can be built with the use of
smaller modules. The modules were constructed out of a single LTM and STM.
2.0
Background
During the latter part of the 20th
century, the study of the brain moved from a peripheral position within both
the biological and psychological sciences to become an interdisciplinary field
called neuroscience that now occupies a central position within each discipline.
This realignment occurred because the biological study of the brain became
incorporated into a common framework with cell and molecular biology on the one
side and with psychology on the other. In recent years, neuroscientists and
cognitive psychologists have recognized many important distinctions of
different sorts of memory. There is a lot of speculations to how the memory is
constructed and what functions it has. Since the memory is a very integrated
system, it’s hard to test specific parts / properties of it. The mixture of two
disciplines is one of the reasons why there are so many ideas to how the memory
is constructed, and the jungle of terminology surrounding the subject.
The different memory systems
have been distinguished according to several attributes or criteria. Here are
some of the more important differences; The content or kind of information
those systems mediate and store (Episodic / Semantic / Procedural memory) and
how they store and retrieve that information (Explicit / Implicit memory).
Another distinction is the memory’s storage capacity and the duration of the
information storage (LTM / STM).
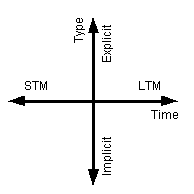
Figure 1 An illustration of how the
properties of memory can viewed to be orthogonal. The horizontal axis represents
the time span of the memories and the vertical axis could be said to represent
awareness of the memories.
LTM can be thought of as a
sturdy memory with almost unlimited capacity. The LTM is thought to reside in
the different receptive areas of the cerebral cortex [11]. A closer description of the receptive areas of the
cerebral cortex is presented in section 2.2. LTM can be seen to be composed of
two different types of memory, declarative memories and nondeclarative
memories. An example of declarative memory is the name of your mother, while
your cycling skill is a nondeclarative memory. The time scale for long-term
memory operations ranges from minutes to years. The time span of a memory depends
on a number of factors. One of the most important factors is the number of
times the memory is presented to you.
The concept of a STM has been
around for a long time. The time scale of short-term memory operations ranges
from less than a second to minutes. It’s an appealing idea that it exists some
sort of temporary memory storage where sensory impressions could temporarily be
stored before they are processed or before they become consolidated into LTM.
Several kinds of STM have been described, again mainly on the basis of
storage-time distinctions and phenomenal or neuropsychological data. The
shortest of STM would be iconic memory [12], which has the capacity to retain a visual image for
up to 1 second after presentation. Echoic memory is used to store sounds, and
has a slightly longer time span then iconic memory. Immediate memory
would last a few seconds longer. Although different STM have been proposed, I
will not deepen the discussion into the subject of different kinds of STM,
instead I will adapt a broader view of the subject.
The definition of STM that
transcends the temporal criterion is working memory. Working memory is a
concept of STM that derives from cognitive psychology [8]. Working memory is thought to be a temporary storage
used in performance of cognitive behavioural tasks, such as reading, problem
solving, and delay tasks (e.g., delayed response and delayed matching to
sample), all of which require the integration of temporally separate items of information.
Baddeley have more recently developed his view of working memory, and he now
states that it constitutes of a phonological loop, a visuospatial sketchpad and
the central executive [8].

Figure 2 A hierarchic view of the
constituents of explicit and implicit memory. Explicit memories, are memories
that you are aware of. Implicit memories are memories you possess, but are not
aware of. Explicit memories can be divided into two categories, episodic and
semantic memories. Episodic memories are whole scenarios. Semantic memories are
lexical memories i.e. words. A form of implicit memory is procedural memory. As
mentioned earlier, the group of implicit memories are memories you are not
aware of i.e. the skill of cycling.
Explicit (or declarative) memory is the memory
of events and facts; it is what is commonly understood as personal memory. One
part of it contains the temporally and spatially encoded events of the
subject’s life for which reason it has alternately been called episodic
memory [13, 14]. Another part contains the knowledge of facts that
are no longer ascribable to any particular occasion in life; they are facts
that, through single or repeated encounters, the subject has come to categorize
as concepts, abstractions, and evidence of reality, without necessarily
remembering when or where he or she acquired it. This is what Tulving has
called semantic memory [14].
Implicit (or nondeclarative)
memory, the counterpart of declarative memory, is a somewhat difficult concept
to grasp. It can be viewed as the memory for development of motor skills
although it encompasses a wide variety of skills and mental operations. Cohen
and Squire called this type of memory procedural memory [15]. Implicit memory can also be viewed as the influence
of recent experiences on behaviour, even though the recent experiences are not
explicitly remembered. For example, if you have been reading the newspaper
while ignoring a television talk show, you may not explicitly remember any of
the words that they used in the talk show. But in a later discussion, you will
more likely use the words that they used in the talk show. Psychologists call this phenomenon priming,
because hearing certain words “primes” you to use them yourself.
The nervous system consists of
the central nervous system and the peripheral nervous system. The central
nervous system (CSN) is the spinal cord and the brain, which in turn include a
great many substructures. The peripheral nervous system (PNS) has two
divisions: the somatic nervous system, which consists of the nerves that convey
messages from the sense organs to the CNS and from the CNS to the muscles and
glands, and the autonomic nervous system, a set of neurons that control the
heart, the intestines and other organs.
The brain is the major component
of the nervous system and it is a complex piece of “hardware”. Weighing
approximately 1.4 kilogram in an adult human, it consists of more than 1010
neurons and approximately 6*1013 connections between these neurons [16]. The struggle to understand the brain has been made
easier because of the pioneering work of Ramón y Cajál [17], who introduced the idea of neurons as structural
constituents of the brain. I will now make some comparisons that are far from
valid but although quite illustrative. Typically, neurons are five to six
orders of magnitude slower than silicon logic gates; events in a silicon chip
happen in the nanosecond (10-9 s) range, whereas neural events
happen in the millisecond (10-3 s) range. However, the brain makes
up for the relatively slow rate of operation of a neuron by having a truly staggering
number of neurons with massive interconnections between them. Although the
brain constitutes an incredibly large number of neurons, it’s still very energy
efficient. The brain use approximately 10-16 joules per operation
per second, whereas the corresponding value for the computers in use today is
about 10-6 joules per operation per second [18]. If one makes the assumption that the brain consumes
400 kg-calories/24h, the brain has an effect of 20 watts, which is equal to a
modern processor.
What sets neurons apart from
other cells are their shape and their ability to convey electrical signals. The
anatomy of a neuron can be divided into three major components: the soma (cell
body), dendrites and an axon. The soma
contains a nucleus, mitochondria, ribosomes and the other structures typical of
animal cells. Neurons come in a wide variety of shapes and sizes in different
parts of the brain. The pyramidal cell is one of the most common types of
cortical neurons. The typical pyramidal cell can receive more than 10,000
synaptic contacts, and it can project onto thousands of target cells. Axons are
the transmission lines from the soma to the synapse, and dendrites are the
transmission lines from the synapses to the soma. These two types of cell
filaments are often distinguished on a morphological ground. An axon often has
few branches and greater length, whereas a dendrite has more branches and
shorter length. There are some exceptions to this view. Some dendrites contain
dendritic spines where specialized axons can attach [19].
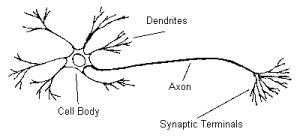
Figure 3 A pyramidal
cell, which is the most common type of nerve cell in the cerebral cortex. The
pyramidal cell is here depicted with only its most important filaments, the
dendrites, the axon with its synaptic terminals, and the cell body. [Figure 3]
Synapses are elementary
structural and functional units that mediate the interactions between neurons.
The most common kind of synapse is a chemical synapse. A presynaptic process
liberates a transmitter substance that diffuses across the synaptic junction
between neurons and then acts on a postsynaptic process. Thus a synapse
converts a presynaptic electrical signal into a chemical signal and then back
again into a postsynaptic electrical signal. It is assumed that a synapse is a
simple connection that can impose excitation or inhibition, but not both on the
receptive neuron. It is established that the synapses can store information
about how easily signals should be able to pass through. The process that
account for this ability is LTP (Long Term Potentiation). In the case of
inhibitory synapses there is a similar process called LTD (Long Term
Depression) [1].
The majority of neurons encode
their outputs as a series of brief voltage pulses. These pulses, commonly known
as actionpotentials or spikes, originate at or close to the soma (cell body) of
neurons and then propagate across the individual neurons at constant velocity
and amplitude. The reasons for the use of actionpotentials for communication
among neurons are based on the physics of axons. The transportation of the
actionpotentials is an active process. The axon is equipped with ion-pumps,
that actively transport K+, Na2+, Cl-, ions in
and out through the axon’s cell membrane. The active transportation of
actionpotentials is necessary when the axons span great distances otherwise the
actionpotentials would be too reduced. If the actionpotentials are too reduced
when they reach the end of the synapses, they are not able to initiate the
release of transmitter substances. The myelin or fat that surrounds the axons
lessen the reduction of the actionpotentials.
The surface of the forebrain
consists of two cerebral hemispheres, one on the left side and one on the
right, that surround all the other forebrain structures. Each hemisphere is
organized to receive sensory information, mostly from the contralateral side of
the body, and to control muscles, mostly on the contralateral side, through
axons to the spinal cord and cranial nerve nuclei. The cellular layers on the
outer surface of the cerebral hemispheres form grey matter known as the
cerebral cortex. Large numbers of axons extend inward from the cortex, forming
the white matter of the cerebral hemispheres. Neurons in each hemisphere
communicate with neurons in the corresponding part of the other hemisphere
through the coreus callosum, a large bundle of axons.
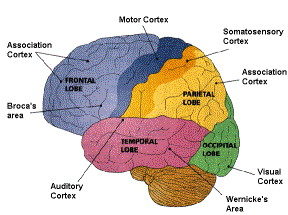
Figure 4 The cerebral cortex of a human
brain. In the picture the cortex has been divided into it’s major functional
areas. There are descriptions of the cortex, where the cortex is divided into
50 functional areas or more. [Figure 4]
The cerebral cortex has a very
versatile functionality. At a glance, the cortex seems to be structurally very
uniform. This suggests that the functionality of the cortex is very general,
but we know that different areas of the cortex handle specific tasks. This is
supported by the fact that microscopic structure of the cells of the cerebral
cortex varies substantially from one cortical area to another. The differences
in appearance relate to differences in the connections, and hence the function.
Much research has been directed toward understanding the relationship between
structure and function. The sensory and motor cortical areas have been found to
have a hierarchy order. In the case of the sensory cortical areas, there are
lots of connections from the higher order sensory areas to the prefrontal
cortex. In the case of the motor cortical areas, there are lots of connections
leading from the prefrontal cortical area to the higher order motor cortical
areas [20].

Figure 5 The cerebral cortex is divided
into to six layers. Layer 2&3 is often considered as a single layer. There
is also a thought of a division of the cortex into columns vertical to the
layers of cortex. The existence of these vertical modules is highly debated.
[Figure 5]
In humans and most other
mammals, the cerebral cortex contains up to six distinct laminas, layers of
cell bodies that are parallel to the surface of the cortex. Layers 2 and 3 are
usually seen as one layer. Most of the incoming signals arrive in layer 4. The
neurons in layer 4 sends most of their output up to layers 2 & 3. Outgoing signals
leaves from layers 5 and 6. In the sensory cortical areas the cells or neurons
with similar interests tend to be vertically arrayed in the cortex, forming
cylinders known as cortical columns. The small structures, called mini-columns,
are about 30 mm
in diameter. These columns are summed
up into larger structures called hypercolumns that are about 0.4 -1.0 mm. In
the artificial neural network used to run the simulations described later on,
there will be a similar concept to the hypercolumns. Outside the sensory areas
the structures of the columns are less distinct. Each column in a hypercolumn
can be seen to perform a small and specific piece of the work that is preformed
by a hypercolumn. Within a hypercolumn the communication between the columns constituting
the hypercolumn, is very intensive [21].
Neural networks are very
interesting, because they work in a completely different way than a
conventional digital computer does. Neural networks process information, using
a vast number of non-linear computational units. This means that the
computations are done in a non-linear and highly parallel manner. A
conventional computer, based on the von Neumann machine, often only uses one
computational unit, hence process the information in a sequential manner. It is
often said that neural networks are superior to the standard von Neumann
machines. This isn’t true, but it’s a fact that neural networks and von Neumann
machines are good at different forms of computations [22].
The neural networks used in this
thesis were implemented on regular desktop computers. This is usually the case,
since it’s much easier to construct an implementation in software than in
hardware. A hardware implementation of a neural network is more resource
efficient.
An associative memory is a
memory that stores its inputs without labelling them (memories aren’t given an
address). To recall a memory you need to present the associative memory with an
input similar to the memory you want to retrieve. There are two types of
associative memories, auto-associative memories (which are sometimes
also referred to as content addressable memories) and hetro-associative
memories. When fragmented pattern is presented to an auto-associative
memory, the memory tries to complete the pattern. If a fragmented pattern is
presented to a hetro-associative memory, the memory tries to associate the
presented pattern to another pattern. Note that all the associations are
learned in advance [23].
The basic idea behind an
auto-associative memory is very simple. Each memory is represented by a
pattern. A pattern is a vector containing N binary values corresponding to the
states of the N neurons. When an auto-associative memory has been trained with
a set of P patterns { xm } and then presented with a new pattern xP+1,
the auto-associative memory will respond with producing whichever one of the
stored patterns most closely resembles xP+1. This could of course be
done with a conventional computer program that computes the Hamming distance
between pattern xP+1 and each of the P stored patterns. The Hamming
distance between two binary vectors is the number of bits that are different in
the two vectors. But if the patterns are large, and very many (these two
attributes usually come together), the auto-associative memory with its highly
parallel structure will be immensely faster then the conventional computer
program. An example of application is image recognition. Imagine that you
receive a very noisy image of your house, if this image previously has been
stored in the auto-associative memory, the memory will produce a reconstruction
of the image.
Associative memories have more
nice features then just noise removal. Associative memories have the very
import ability to generalize. This makes it possible for associative memories
to handle situations where they are presented with memories never before
encountered. Another side of generalization is categorization of memories. This
is also a feature handled by associative memories. Categorization means that
similar memories are stored as one memory. The common features of the memories,
stored in the same category, are stored robustly and are easy to retrieve. The
individual details of each memory in the category leave only a minor trace in
the memory. When a memory is retrieved from the category, it is very likely to
posses the details of the most recently stored memory.
The workings of the associative
memory are usually explained by an energy abstraction. In this abstraction, the
memories stored in the associative memory, constructs an energy landscape. The
energy landscape has as many dimension as the stored memories have attributes.
This often means that the energy landscape has a high number of dimensions. The
energy landscape in figure 6 only has two dimensions and thus the memories
stored, in the corresponding associative memory, only have two attributes. Each
learned memory creates a local minimum in the energy landscape. These local
minimums are called attractors. In this concept, the input to the associative
memory is a position in the energy landscape, where information is stored as a
basin in the energy landscape. The retrieval of a memory can be seen as a
search for a local minimum in the energy landscape. The starting point for this
search is the input, which is similar to the memory that is going to be
retrieved. Although this view can be quite elusive, since we are talking about
a high dimensional space, it nonetheless gives an illustrative view of the way
an associative memory works.
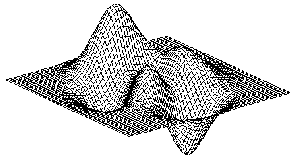
Figure 6 An illustration of the energy
landscape that is produced by the associative memory. The basins, the lowest
points in this energy landscape, are called attractors. The attractors could be
said to constitute the memories in an associative memory. These types of
networks are referred to as attractor networks.
There are several ways an
associative memory can be constructed. The most common method is to use the
Hopfield model to construct an associative memory. In this thesis I will use an
advanced version of the Hopfield model, based on the laws of probabilities, to
construct associative memories.
The idea behind the Hopfield
model is largely based on Donald Hebb’s well known work: Assume that we have a
set of neurons, which are connected to each other through connection weights
(representing synapses) [24]. In the discrete Hopfield model, the neurons can
either be active or non-active. When the neurons are stimulated with a pattern
of activity, correlated activity causes connection weights between them to
grow, strengthening their connections. This makes it easier for neurons that in
the past have been associated to activate each other. If the network is trained
with a pattern, and then presented with a partial pattern that fits the learned
pattern it will stimulate the remaining neurons of the pattern to become
active, completing it. If two neurons are anti-correlated (one neuron is active
while the other neuron is not) the connection weights between them are weakened
or become inhibitory. This form of learning is called Hebbian learning, and is
one of the most used non-supervised forms of learning in neural networks.
The Hopfield network consists of
a set of neurons and a corresponding set of unit delays, forming a
multiple-loop feedback system [9]. If N is the number of neurons in the network, the
number of feedback loops is equal to N2-N. The ”–N” expression
represents the exclusion of self-feedback. Basically, the output of each neuron
is fed back, via a unit delay element, to each of the other neurons in the
network. Note that the neurons don’t have self-feedback. The reason to this is
that self-feedback would create a static network, which in turn means a
non-functioning memory.
Each feedback-loop in the
Hopfield network is associated with a weight, wij. Since we had N2-N
feedback loops, we will have N2-N weights. Imagine that we have P
patterns, where each pattern, xm, is a vector containing the values 1 or –1. Then
a weight matrix can be constructed in the following manner:
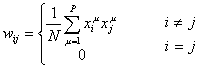
where m is index within a set of
patterns, P is the number of patterns, and N is the number of units in a
pattern (N is the size of the vectors in the set { xm
}). The patterns represent the activation of the neurons. The neurons can be in
the states oi Î ±1.
To recall a pattern (of
activation), oi, in this network we can use the following update
rule:
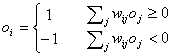
If the underlying network is
recurrent the process of recollection is iterative. This iterative process,
where the instable and noisy memory becomes stable and clear, is called relaxation.
Since the network will have a
symmetric weight matrix, wij, its possible to define a energy
function called Lyapunov function [25]. The Lyapunov function is a finite-valued function
that always decreases as the network changes states during relaxation.
According to Lyapunov’s Theorem 1, the function will have a minimum somewhere
in the energy landscape, which means the dynamics must end up in an attractor.
The Lyapunov function is for a pattern x defined by:
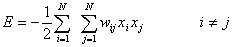
The Hopfield model constitutes a
very simple and appealing way to create an associative memory. The model have a
problem called catastrophic forgetting.
Catastrophic forgetting occurs when the Hopfield network is loaded with
too many patterns. It can be said to occur when there are to many basins in the
energy landscape. If the network is loaded with to many patterns, errors in the
recalled patterns will be very severe. The storage capacity of the Hopfield
network is approximately 0.14N patterns, where N is the numbers of neurons in
the network [25].
The Hopfield model can also be
made continuous. The model is then described by a system of non-linear first-order
differential equations. These equations represent a trajectory in state space,
which seeks out the minima of the energy (Lyapunov) function E and comes to an
asymptotic stop at such fixed points in analogy with discrete Hopfield model
presented.
The standard correlation based
learning rule used in the Hopfield model, suffers from catastrophic forgetting.
To cope with this situation Nadal, Toulouse and Changeaux [26] proposed a so-called marginalist-learning
paradigm where the acquisition intensity is tuned to the present level of cross
talk “noise” from other patterns. This makes the most recently learned pattern
the most stable. New patterns are stored on top of older ones, which are
gradually overwritten and become inaccessible, a so-called “palimpsest memory”.
This system retains the capacity to learn at the price of forgetfulness.
Another smoothly forgetting
learning scheme is learning with in bounds, where the synaptic weights wij are bounded –A £ wij £ A. This learning scheme
was proposed by Hopfield [9]. The learning rule for training patterns xn
is
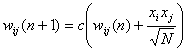
where c is a
clipping function
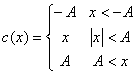
The optimal capacity 0.05N is
reached for A »
0.4 [27]. For high values of A, catastrophic forgetting
occurs, for low values the network remembers only the most recent pattern. This
implies a decrease in storage capacity from 0.14N of the standard Hopfield
model. Total capacity has been sacrificed for long-term stability.
As previously discussed, there
are several approaches to creating a memory in a neural network context. This
thesis uses associative memories with, palimpsest properties and a structure
with hypercolumns based on a Bayesian attractor network with incremental
learning. This memory model is used because it is a good model of the
structures in the cerebral cortex and at the same time comparably simple. The
model also makes sense from a statistical viewpoint.
The artificial neural network
with hypercolumns and incremental, Bayesian learning developed by Sandberg, et
al. [10] is a development of the original Bayesian artificial
neural network model developed by Lansner et al. [28-30] , which was developed to be used with one-layer
recurrent networks.
The Bayesian learning method is
a learning rule intended for units that sum their inputs multiplied by weights
and use that sum to determine, by using a non-linear function, their output
(activation). This is much like many other algorithms for artificial neural
networks. The weights in a Bayesian network are set in accordance with rules
derived from Bayes’ expressions concerning conditional probabilities. This
means that the unit activation can be equated with the confidence in various
features. The rule is local, i.e. it only uses data readily available at either
end of a connection. The algorithm allows, in an easy way, to adjust the time
span over which statistical data is collected. The time span is adjusted by a
single variable, often called a.
Regulating the value of a
, and hence the time span for collecting statistical data, the plasticity of
the network is regulated.
The Bayesian learning rule can
be extended to handle continuous valued attributes. This have been done by
Holst and Lansner [30], using an extended network capable of handling graded
inputs, i.e. probability distributions given as input, and mixture models.
To deal with correlations
between units that cause biases in the posterior probability estimates,
hypercolumns were introduced [29]. A hypercolumn, named in analogy with cortical
hypercolumns [31], is a module of units that represent all possible
combinations of values of some primary features and hence provide a
anti-correlated representation of the network input.
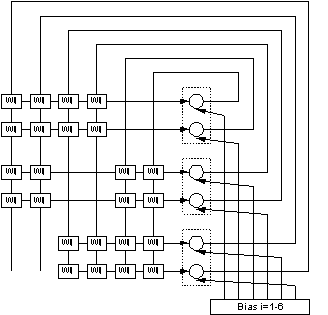
Figure 7 Here is a small recurrent
neural network with the 6 neurons divided into three hypercolumns. Note that
there are no recurrent connections within each hypercolumn. With some
imagination it can also be seen how the weights wij form a
matrix.
I am now going to present a
continuous, incremental Bayesian learning rule with palimpsest memory
properties. The forgetfulness can
conveniently be regulated by the time constant of the running averages. This
implies that we easily can construct a STM or LTM memory with this learning
rule.
Bayesian Confidence Propagation
Neural Networks (BCPNN) are based on Hebbian learning and derived from Bayes
theorem for conditional probabilities:
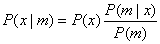
where m is an attribute value of
certain class x. The purpose of calculating the probabilities of the observed
attributes for each class is to make as few classification errors as possible.
The reason we want to use Bayes theorem is that it is often impossible to make
a good estimate of P(x|m) directly out of the training data set. On the other
hand, a good estimate of P(m|x) is often possible to achieve. Next we will see
how this can be implemented in a neural network context.
The input to the network is a
binary vector, x. The vector x is composed of the smaller vectors
x1, x2,…xN. Each of these sections x1,
x2,…xN are representing the input to a hypercolumn. This
means that the input space, which represents all possible inputs to the
network, can be written as X = X1, X2,…XN
Each variable Xi can
take on a set of Mi different values. This means xi will
be composed of Mi binary component attributes xii’ (the
i’ th possible state of the i th attribute of xi) with a normalised
total probability
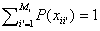
From the input, x, we
want to estimate the probability of a class or set of attributes y. (The class y is the output of
the network and the input, x, is seen as an attribute.) The vector y
has the same structure as the vector x. If
we condition on X (where unknown attributes retain their prior distributions)
and assume the attributes xi, to be both independent, P(x) = P(x1)P(x2)…P(xN), and
conditionally independent, P(x|y) = P(x1| y)P(x2| y)…P(xN| y),
we get:
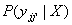

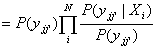
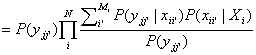

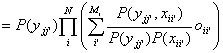
where oii’
= P(xii’|Xi).
Since y can just be regarded as another random variable it can be
included among the attributes xi and there is no reason to
distinguish the case of calculating yjj’ from calculating xii’.
If X represents known or estimated information, we want to create a
neural network which calculates P(y),
from the given information. If we take the logarithm of the above formula we
get
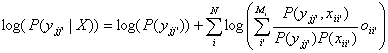 (1)
(1)
Now, let the input X(t) to the network be viewed as a
stochastic process X(t,×) in
continuous time. Let Xii’(t) be component ii’ of X(t), the observed input. Then we can
define Pii’(t)=P{X ii’(t)=1} and P ii’jj’(t)=P{X
ii’(t)=1, X jj’(t)=1}. Equation (1) becomes:
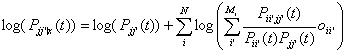 (2)
(2)
Given the information
{X(t’),t’<t} we now want to estimate Pii’(t) and P ii’jj’(t).
This can be done by using current unit activity oii’(n) at time n with the following two estimators
where t
is a suitable time constant.
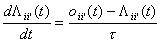 (3)
(3)
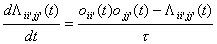 (4)
(4)
The estimator in equation (3)
estimates the probability of a single neuron to become active per time unit.
The estimator in equation (4) estimates the probability for two neurons to
simultaneously be active. L is the estimated probability per time unit or rate
estimated probability. This means that L is estimated from a
subset of the events that has occurred, whereas P is estimated from all events
that has occurred. The rate estimator explains the palimpsest property of the
learning rule.
These estimates can be combined
into a connection weight, which is updated over time. The bias can also easily
be stated:
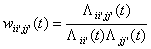 (5)
(5)
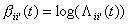 (6)
(6)
The base for the logarithms is
irrelevant, but for performance reasons the natural logarithm is often the best
choice. Logarithms with other bases are often derived from the computation of
the natural logarithm.
The usual
equation for neural network activation is
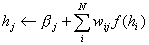 (7)
(7)
where hj is the
support value of unit j, bj is its bias, wij the weight from
i to j and f(hi) the output of unit i calculated using the transfer
function f. The output f(hi) equals oii’ in equation (1)
and (2). In the basic Hopfield model the activation function, f(), is, as we
earlier saw, a step function.
The form in equation (2) is
slightly more involved then (7), and has to be implemented as a pi-s
neural network or approximated [29,
30]. The activation equation in the learning rule is
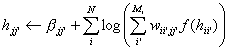 (8)
(8)
Comparing
terms in equation (8) and (2) we make the identifications
 (9)
(9)
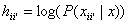 (10)
(10)
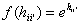 (11)
(11)
P(xii’|x) = oii’
= f(hii’) can be identified as the output of unit ii’, the
probability that event xii’ has occurred or an inference that it has
occurred. Since inferences are uncertain, it is reasonable to allow values
between zero and one, corresponding to different levels of confidence in xii’.
Since the independence
assumption is often only approximately fulfilled and we deal with
approximations of probability, it is necessary to normalise the output within
each hypercolumn:
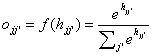 (12)
(12)
The network is used with an
encoding mode where the weights are set and a retrieval mode where inferences
are made. Input to the network is introduced by setting the activations of the
relevant units (representing known events or features). As the network is
updated the activation spreads, creating a posterior inferences of the
likelihood of other features.
As we discussed earlier,
networks with update rules like equation (7) and symmetric weight matrices an
energy function can be defined, and convergence to a fixed point is assured [27]. In this case this does not strictly apply, but for
activation patterns leaving only one nonzero unit in each hypercolumn, it does
apply. In practice it almost always converges, even though there is no input.
In the absence of any
information, there is risk for underflow in the calculations. Therefore we
introduce a basic low rate l0. In the absence of signals Lii’(t)
and Ljj’(t)
now converges towards l0 and Lii’jj’
towards l20,
producing wii’jj’(t) = 1 for large t (corresponding to uncoupled
units). The smallest possible weight value if the state variables are
initialised to l0
and l20
respectively is 4l20,
and the smallest possible bias log (l0). The upper bound on the weights becomes 1/l0.
This learning rule is hence a form of learning within bounds, although in practice
the magnitude of the weights rarely comes close to the bounds.
The learning rule (equations
(3)-(6)) of the preceding section can be used in an attractor network similar
to the Hopfield model by combining them with an update rule similar to
equations (8)-(12). The activities of the units can then be updated using a
relaxation scheme (for example by sequentially changing the units with the
largest discrepancies between their activity and their support from other
units). One could also use a random or synchronous updating similar to an
ordinary attractor neural network, moving it towards a more consistent state.
This latter approach is used here. The continuous time version of the update
and learning rule takes the following form (the discrete version is just a
discreteisation of the continuous version using Euler’s method):
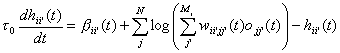 (13)
(13)
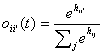 (14)
(14)
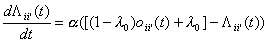 (15)
(15)
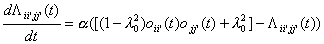 (16)
(16)
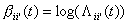 (17)
(17)
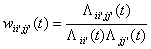 (18)
(18)
where t0 is the time
constant of change in unit state. a = 1/t is the inverse of the learning time constant; it is a
more convenient parameter than t. By setting a temporarily to zero the network activity can change
with no corresponding weight changes, for example during retrieval mode.
The use of hypercolumns in the
model presented implies that there will be no recurrent connections within a
hypercolumn of the network. Recurrent connections within a hypercolumn are
fully anti-correlated. The self-recurrent connection is fully correlated. Thus
the weights connecting the neurons within a hypercolumn would either be set to
their minimum or their maximum value.
Each neuron in the network will
have a bias that is derived from the basic set of recurrent connections.
Connections projected from other populations of neurons will not add any bias to
the receiving neurons, although it would make sense from a mathematical point
of view to include the bias in the projection.
Auto-associative memories based
on artificial neural attractor networks, like for example early binary
associative memories and the more recent Hopfield net, have been proposed as
models for biological associative memory [9,
32]. They can be regarded as formalisations of Donald
Hebb’s original ideas of synaptic plasticity and emerging cell assemblies. In
this view each neuron in the artificial neural network is thought to equal a
single nerve cell in the biological neural network. In figure 8 is an
illustration of an artificial neuron. With some imagination it is possible to
see the similarities with a nerve cell.
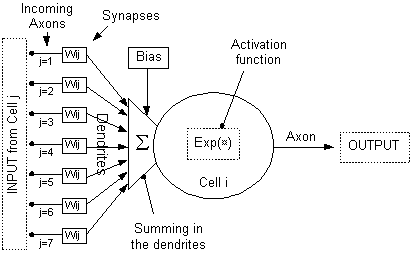

Figure 8 Depicted
here is an artificial neuron, and its functions. Some parallels to a biological
neuron are inferred in the figure. Note that the output is conveyed to several
other neurons.
Each connection weight, wij,
in figure 8 can be interpreted as a synaptic connection between two neurons. In
figure 9 one of these connections is depicted in more detail.
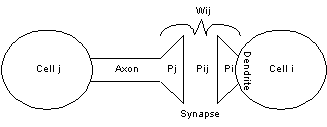
Figure 9 This figure shows a single
synaptic connection between two neurons in our artificial neural network. The
values of Li (=Pi,
Pj), Lij (=Pij),
and wij , derived in equations (15, 16, 18), can be interpreted as
shown in figure. Pj, Pi, Pij, are values
associated with synaptic terminals, synaps, dendrites ability to convey a
signal from cell j to cell i.
Although the above presented
view of each neuron corresponding to a nerve cell is appealing, it isn’t
realistic. Real neurons aren’t as versatile as our artificial neurons i.e. a
real neuron can’t impose both inhibition and excitation, which is stated by
Dalés law [33]. A better view of the correspondence between our
artificial neurons and real neurons is to think of our artificial neurons as
corresponding to a cortical column of real neurons. In our Bayesian attractor
network we have a structure of hypercolumns, where each hypercolumn is
corresponding to a group of cortical columns.
There are a couple interesting
concepts or functions I hope to find in the simulations of the memory systems
developed in the experiments. These
concepts originate from cognitive psychology.
In a memory that’s going to be
implemented in a decision making system, there’s a need not only to be able to
recall earlier events, but also be able to recall these events with current
situation data. This type of process is usually called short-term variable
binding (STVB) or role filling. To illustrate this concept I will
give an example:
John is visiting his
grandfather Sven. After he has visited his grandfather, John meets his two
friends, Max and Sven. When Max talks about Sven with John, John knows that the
Sven, Max is talking about isn’t his grandfather.
To achieve STVB in a system, the
system will of course need a LTM, and also some sort of STM that can
accommodate the temporary bindings. One of the main focuses of this thesis will
be to investigate how STVB can be achieved.
The chunking process is a
specialisation of the memory, which allows it to more effectively remember
certain things. The chunking learning process recruits a new idea to represent
each thought, and strengthens associations in both directions between the new
chunk idea and its constituents. Thus, the inventory of ideas in the mind does
not remain constant over time, but rather increases due to chunking. The
representation of a chunk is constructed out of its constituents. An example of
chunking is how the set { 1 2 3 } is remembered. The set can be remembered as
1, 2, and 3. The chunked version of the set is remembered as the number 123.
There are two primary reasons
for chunking: First, chunking helps us to overcome the limited attention span
of thought by permitting us to represent thoughts of arbitrary complexity of
constituent structure by a single (chunk) idea. Second, chunking permits us to
have associations to and from a chunk idea that are different from the
associations to and from its constituent ideas. This is very important for
minimizing associative interference.
In this thesis I have studied
how the representation of the short-term memories could be done more effectively.
I have also studied how these, efficient short-term representations associates
to the long-term representations. Although, the work of this thesis does not
directly focus on the chunking process, I thought it was interesting to mention
the similarities between the STM & LTM interaction and chunking.
3.0 Method
Since the simulations in this
thesis are based upon the Bayesian artificial neural network model developed in
[10], I tried to use similar settings and architectures.
In all simulations, the neural networks were first trained on a set of
patterns, and then tested. This means some consideration must be taken before
the artificial neural networks of this thesis can be implemented in a real-time
system.
The LTM was implemented as a
recurrent network consisting of 100 neurons divided into 10 hypercolumns with
10 neurons in each hypercolumn. This configuration of LTM was used throughout
the thesis with no exceptions. As for the STM, there were a couple of different
implementations in respect to the number of neurons and of hypercolumns. The
two most common implementations of the STM used 100 and 30 neurons
respectively. All the networks used, had at least one set of recurrent
connections. As earlier mentioned there were no recurrent connections within
the hypercolumns of the networks, as the internal representation in a
hypercolumn is supposed to be completely anti-correlated.
Almost all systems were
constructed under the assumption that the input to the systems always passed
the LTM before it entered the STM. The output from the system was always
extracted through the LTM. When the systems are used and not only tested, all
input/output is handled by the LTM. In a real-time system, the data presented
to the STM will always be delayed. Since the simulations in this thesis were
not run in real-time, there was no need to be concerned about this delay. The
LTM exerted a disruptive influence on the STM during training. The STM exerted
a disruptive influence on the LTM during operation. In chapter 5 I investigated
these interferences.
The recurrent connection within
a network and the connections between networks are called projections. A
projection does not only represent the physical connection, the concept also
incorporates the connection-weights. Each connection, between two neurons, is
equipped with two weights that represent the correlation between the neurons in
both directions. Since the networks are auto-associative, the weights are equal
in both directions. This does not apply to the connections between two
networks, where hetro-associations may arise. In the models, a matrix
represents the projections. The bias was not included in the projections.
The input to the artificial
neural networks was vectors of binary numbers (0 and 1). These input vectors
were constructed in respect to the hypercolumn structure of the LTM. This meant
that only one out of ten neurons in a hypercolumn structure was activated and
this was always the case. So the whole input of ten hypercolumns only caused
10, out of 100, neurons to be activated. The input could therefore be
considered sparse. The sparseness of the input affect the storage
capacity of the network. If the input is to “dense”, it will affect the storage
capacity negatively. In every run a new set of patterns was generated. The
patterns were generated from a rectangular probability-density function.
In chapter 4 the input always
consists of sets with 100 patterns. In chapter 5 the input always consists of
sets with 50 patterns. Chapter 6
contain experiments with structured data. Therefore the input is sets with
different number of patterns. The number of patterns in an input set does not
affect the STM since it forgets so quickly. The LTM is affected by the size of
the input set. This is more thoroughly explained in 3.2.
As mentioned earlier, the
systems designed in this thesis were not operated in “real-time”. The systems
had a training mode, where the memories were stored in the system. Then, during
operation mode the memories were retrieved. The system design outlined in
figure 10 was the most frequently used design. In biological memory systems the
theta rhythm may control the switch between training and operation mode of the
memory network.
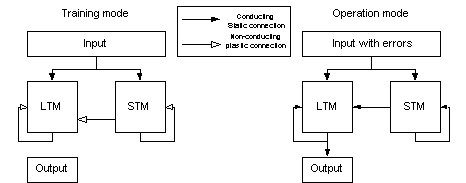
Figure 10 In all
simulations, the artificial neural networks were first put in a training
mode and were trained with a set of patterns. When the training phase was completed, the networks were put in operation
mode, and tested. Note that non-conducting static projections are not
depicted in the figure (training mode).
The differential equations 13,
15 and 16 have been solved with Euler’s method. The time-step was chosen to
h=0.1, and the integrations lasted for 1 unit of time. (This meant that 10
steps were taken during integration from 0 to 1.) It often took much longer
time than one time unit to train or retrieve a pattern properly. In the case of training, a strong memory of
a single pattern was achieved trough repeated presentation of the pattern. The
relaxation process that occurs during operation mode was almost never fully
completed. (Fully completed mean that no more changes would occur if the process
was extended.)
During the training mode the
equations (15) and (16) were solved for each network in the system. Equations
(17) and (18) were then used to compute the bias and the projections for the
networks. In the case of projections between two networks, the same equations
were applied with the exception of equation (17). The bias was chosen not to be
included in the projections between networks. This could of course be
discussed. A biological interpretation of this is that the whole dendritic tree
of the synapse is given the same bias value. This means that synapses close to
the soma are not given any priority. In a real neuron, synapses closer to the
soma generate a stronger signal then synapses further out in the dendritic tree
[34]. Mathematically it also makes sense to incorporate
the bias values into the projection, even though this was not done here.
The three main parameters that
controlled the network during training mode were the value of a, the
number of patterns and the time spent training each pattern.
The main interest of this thesis
was not the retrieval-performance of the networks. The main interest was to
prove that networks with different time-dynamics could be used in the same
system. However, retrieval-performance was of great importance when different
designs were investigated, to rate how good the designs were.
To initiate the retrieval of
patterns (memories) the networks were usually presented with a copy of the learned
pattern with 2 errors. (The content of two of the hypercolumns were altered.)
In the figures describing the systems, this type of input is denoted as “Input
with errors”. In some experiments of chapter 5 and 6 the networks were
presented with only a few of the hypercolumns of the learned patterns. The
hypercolumns that were not presented to the network were filled with
zeros.
The plots over single networks
were constructed from 50 runs. In the plots of several networks, each data
point was often constructed from 20 runs. The data presented in the tables were
accumulated from 100 runs of the networks.
During testing, the networks
were put in operation mode. The equation (13) and (14) were used in order to
perform the relaxation. The relaxation process was always one time unit long.
When a network had a projection from another network, equation (13) was
replaced with equation (19). Equation (19) introduces the constant gain factor g.
The value of g varied around 1. The purpose of g was to introduce an instrument
to control the influence that connected networks imposed on each other. The
direction of the projections that g applied to was denoted, i.e. gSTM®LTM (In this case the connection from the
STM to the LTM is scaled with g.)
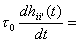
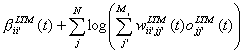
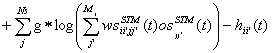 (19)
(19)
Here wsii’jj’ denoted
the connection-weights and osjj’ the activity pattern of the sending
neurons. Ns is the number neurons in the sending network.
Successful retrieval was defined
as the fraction of patterns, which were correctly recalled after relaxation to
a tolerance of 0.85 overlap. In the normal case where the input consisted of 10
hypercolumns, a recalled pattern was only allowed to differ in one hypercolumn
from the original pattern to be classified as correct. The retrieval ratio of
the system was often plotted as a continuous line. The retrieval-ratio of the
subsystems were often also plotted, i.e. the LTM (plotted as a dotted line) and
the STM (plotted as a dash-dotted line).
In the neural network model at
hand there are several parameters that can be chosen more or less arbitrarily.
As mentioned earlier, the choice of these parameters is consistent with [10]. In this section the default values of the parameters
are listed. These default values are common among many of the simulations. When
new constants are introduced or when the default values are altered it has been
mentioned it in the text.
In all simulations, the value
0.001 is used for l0.
l0
can be seen as the background noise in the neurons. l0
also have implications on the maximum excitation that can be conveyed from one
neuron to another.
The experiments of chapter 4
were run with 100 patterns. The capacity for an optimal trained LTM with 100
neurons is about 60 patterns. The LTM in the experiments of chapter 4 had a set to a =
0.0005. This low value of a
implied that the LTM of chapter 4 could not form properly “deep” attractors
after training for 1 unit of time. These two conditions generated a situation
where the memories stored in the LTM had a small chance of correct retrieval.
Although all trained memories left some sort of trace in the LTM. Chapter 4
investigated the possibility of using a STM to extract those memory traces.
Chapter 5 & 6 contains
experiments where the systems where presented with 50 patterns and the LTM was
run with a
= 0.005. This setting of a
allowed the LTM to learn all 50 patterns.
The STM networks in this thesis
always had the value of a
set a
= 0.5. I tried to choose a
in such a way that the STM remembered the 10 most recent patterns presented to
it. This means that the STM wasn’t affected if the number of patterns presented
to it was 50 or 100. Contrary to the LTM of chapter 4, there were no memory traces
stored in the STM of the first patterns in the training set.
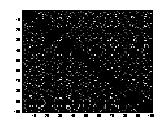
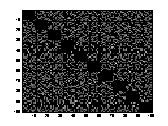
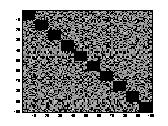
A1 B1 C1
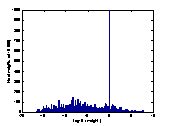
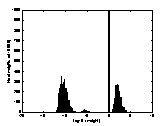
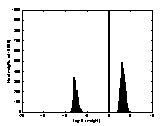
A2 B2 C2
Figure 11 Three connection-weight /
projection matrices. .A1 is from a STM. B1 is from a LTM of chapter 5 & 6.
C1 is from a LTM of chapter 4. The strength of the connection-weights is
colour-coded in A1, B1 and C1 between the logarithmic values 0 and 5. The
brighter a dot is, the stronger the connection. The diagonals of the matrices
all have black squares, showing the absence of connections within a
hypercolumn. A2, B2, C2 are the
corresponding distributions of the connection-weights. The vertical line seen
in A2, B2, C2 represents the 1000 self-recurrent connection-weights that have
been deleted.
There was a difference in the
way the projections were set up in a network with a large value of a compared to how the
projections were set up in a network with a small value of a. In a net trained with a
small value of a
(LTM) I found that the distribution of the inhibitory and excitatory weights
was very even and distinct. In figure B2 and C2, one can see that the
connection-weights are either set to be inhibitory or excitatory. There are not
many connection-weights with a value between the two groups of inhibitory and
excitatory connection-weights. While in a network trained with a large value of
a
(STM), the values of the connection-weights were evenly distributed between
inhibitory and excitatory connection-weights.
When I coupled a STM to a LTM,
the STM memory had an interfering effect on the neurons in the LTM. This meant
that the LTM had a smaller probability to relax to the correct pattern. To
prohibit this impairment of the LTM, I introduced a gain constant, gSTM®LTM,
between the STM and the LTM (equation (19)). In the systems that were trained
with 100 patterns the value of gSTM®LTM
was set to 0.03 and in the systems trained with 50 patterns the gSTM®LTM
was set to 0.1. These values were derived from trial and error processes and
they seemed to give the STM a reasonable influence on the LTM.
4.0 Network
structures
Chapter 4 investigate the basic
concepts of connected networks. In the first part of this chapter, the
importance of having recurrent connections with different plasticity kept in
different networks (having a separate STM and LTM) is studied. Then, plastic
connections are studied. It is also studied how plastic connections can be
constructed and used.
The systems in the experiments
of this chapter were trained with sets of 100 patterns. The LTM were trained
with a set
to 0.0005. The LTM had a poor retrieval-ratio of about 0.3. The choice of a meant that all patterns
in the training set were remembered, but very poorly.
The information in a neural
network is stored in the projections. Two systems were studied here. The two
systems had an equal number of connections, but different number of neurons. N
is equal to 100.
The first system, a network of N
neurons had two projections with a total of 2N2-20N connections.
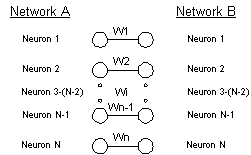
Figure 12 The networks A and B are connected
with one-to-one connections. The connection-weights, wi, were
usually set to a value around 10.
The second system had two
separate networks, with 2N neurons and 2N2-19N connections. The
neurons between these two networks were connected one-to-one projection. This
meant that all elements, except the diagonal elements, in the projection matrix
were set to 1.
The question was, which of these
two systems had the best design. The systems used approximately the same amount
of connections. Since memories are stored in the connections, this comparison
seemed motivated. (Chapter 5 describes how the design of the second system is
made more effective.)
Naturally, a neuron takes much
more space and uses much more resources than a connection between two neurons.
This means that if the number of neurons in a network can be minimized at the
expense of more connections in the network, it is a good thing. The system, in
this experiment, used few neurons and a moderate number of connections.
Real synapses may posses both
low and high plasticity properties. In this experiment the two projections with
different plasticity can be seen to form a single projection that have both low
and high plasticity properties.
The system was based on a LTM. A
“STM” projectionLTM®LTM with high plasticity was added to the system’s
existing “LTM” projectionLTM®LTM with low
plasticity. The high plasticity projection used a set to 0.5 and the low plasticity projection used a set to 0.0005. The bias
values were derived from the training of the low plasticity projection,
“LTM”. The projection with high
plasticity was scaled down with g = 0.03. The value of g
was chosen after evaluating the results of the experiment in section 4.2.1. The
system was trained with equation (19) instead of equation (13).
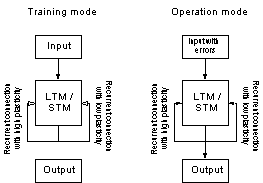
Figure 13 The operation modes of the
system. The bias values were derived from the projection with low plasticity.
The system was constructed with two projections with different plasticity. Each
of the projections, were also treated as individual networks (LTM and
STM).
The retrieval-ratio of the
system is shown as a solid line in figure 14. The systems two projections, with
high and low plasticity were used to create one separate LTM and one separate
STM. The separate retrieval-ratio of the LTM is shown as a dotted line, and the
retrieval-ratio of the STM is shown as a dash-dotted line, in figure 14. The
following text refers to this two (LTM and STM) individualised memories. These
two memories were isolated to provide a base for comparison of
performance.
The retrieval-ratio of the first
80 patterns was slightly lower for the system than for the LTM. The
retrieval-ratio of the last 20 patterns was lower for the system than for the
STM. The system seemed to provide compromise of the retrieval-ratio between the
LTM and STM. Since the high and low plasticity projections in the system were
interacting during the iterative process of relaxation there was a problem with
interference between the two projections. The STM interfered the LTM, during
retrieval of the first 80 patterns. The STM did not have enough influence over
the LTM to control the relaxation process completely, during the last 10
patterns.
It was interesting to see that
the system was able to retrieve patterns 85-90 with slightly higher
retrieval-ratio than the LTM or STM. During the retrieval of these five
patterns the LTM and STM were able to cooperate. This proves that the basic
idea of having several projections with different plasticity in a single system
can be beneficial.
The compromise between a high
retrieval-ratio of the first and the last patterns was controlled by the value
of g.
Adjustments of g could not improve the projections ability to cooperate. These
suggested that this design, with two projections and one population of neurons,
was not optimal.
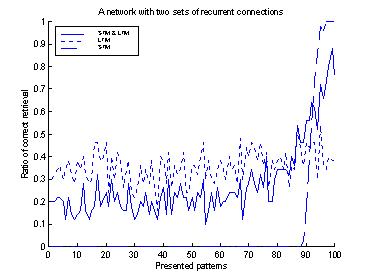
Figure 14 The
retrieval-ratio of the system is plotted as continuous line. The
retrieval-ratio of the LTM is plotted as a dotted line, and the retrieval-ratio
of the STM as a dash-dotted line. Note the increased retrieval-ratio of the
system for the last 10 patterns.
This system basically had the
same two projections as the system in the previous section. The big difference
between the systems was that each of the projections in this system was
projected at a separate group of neurons. The purpose of this experiment was to
determine if it were beneficial too use two networks with different plasticity.
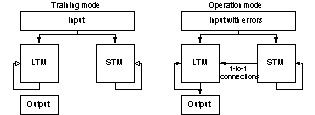
Figure 15 The system has a STM and LTM of
equal size. 1-to-1 connections were used to connect the STM to the LTM. The
input with errors was fed to both the LTM and STM. Output was extracted from
the LTM.
The system was composed a LTM
and STM of equal size. These two memories were connected with a 1-to-1
projectionSTM®LTM. The diagonal elements of the projectionSTM®LTM
were set to 10. This value was derived from a trial and error process seen in
figure 16. When the retrieval-ratio of the system was tested, both of the
networks were fed with input.
The trail and error process to
determine the value of the diagonal elements was performed with ten runs of the
system. For each run, the diagonal elements were set to different values. The
result of these 10 runs is shown in figure 16. The value of the diagonal
elements could have been set to any value between approximately 7-500.
If the diagonal elements, or
weights, had been set to one, there would not have been a connection between
the two memories. Figure 16 shows this fact. When the weights were set to 1
(equal to 0 on the logarithmic x-axel in figure 16) the retrieval-ratio of the
system becomes equal to that of the LTM.
And if the weights had been set to a value smaller than one, there would
have been an inhibitory effect on the neurons in LTM. If the weights had been
set to a value much larger than 500, the system would have shown good
performance on the latest learned patterns, but the system would not have been
able to recall the patterns learned in the beginning of the training set. This
is caused by the strong input from the STM, which makes it impossible for the
LTM to relax into a stable state.
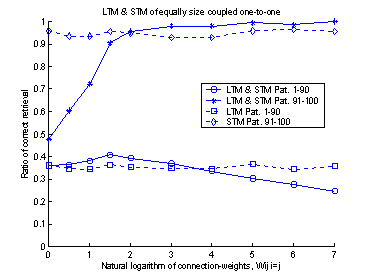
Figure 16 The plot shows 10 runs, with
different values of the connection-weights. Performance is measured separately
for the first 1-90 patterns and the last 91-100 patterns. The dotted lines
represent STM and LTM separately. The solid lines show the performance of the
system.
In figure 17 the retrieval-ratio
of the system is shown. During the first 80 patterns the retrieval-ratio of the
system is equal to that of the LTM. Then, for pattern 80 to 90 the
retrieval-ratio is better than both that of the LTM and STM. During the last 10
patterns the retrieval-ratio is equal to that of the STM.
It was interesting to see that
the cooperation between the two memories (projections in the previous system)
was functioning well. The disruptive influence of the STM on the LTM was almost
negligible. The retrieval-ratio of the last 10 patterns was almost 1. A good retrieval-ratio
of the most recent patterns is necessary when the STM is to be used as a
working memory.
The combination of these facts
proved that the design with two individual networks with different plasticity
was superior to the design of the system in the previous section.
The STM has a strong influence
on the LTM. The STM has the ability to both support and suppress memories in
the LTM with great efficiency. This is an important feature, since it provides
a way to increase the importance of the latest learned patterns. Later on in
this thesis these properties are used to generate useful functions in systems.
Systems with STM are designed to prove the possibility of constructing a
working memory. The STM also provides
the possibility to make reinstatements of the latest memories into the LTM.
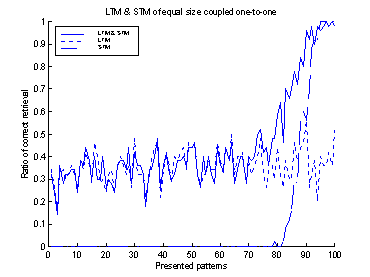
Figure 17 A system was based on a STM and
LTM of equal size. The STM and LTM were connected with one-to-one connections
between the neurons of each network.
The experiments presented in
this section were designed to investigate how a system built upon two networks,
could be connected with plastic projections. Both of the networks, LTM and STM,
were of equal size in all the simulations preformed. Different ideas, of how to
utilise the plastic projections were investigated.
There are many connections
between the neurons in the cortex, especially between neurons that are close
together. It seems very unlikely that these neurons are hardwired and unable to
form new connections or delete old connections. In this experiment, the neurons
of the two networks (STM & LTM) were allowed to form whatever connections
they wanted. As with the recurrent connections, these connections can be made
with different plasticity’s.
The connections between the STM
and the LTM were in this experiment plastic. The projectionSTM®LTM
matrix was no longer diagonal of weights. Instead it was a full matrix of
weights, representing all possible wirings between the neurons of the two networks.
When the size of the STM differs from that of the LTM or when the pattern
representation in the STM differs from that in the LTM, there is a need for a
plastic projection. The plastic projections were trained with the same Bayesian
learning rule that was used to train the networks recurrent projections. The
projectionSTM and the projectionSTM®LTM
were trained with a set
to 0.5. The system’s training and operation mode is seen in figure 18. The
projectionSTM®LTM was scaled down with gSTM®LTM
= 0.03.
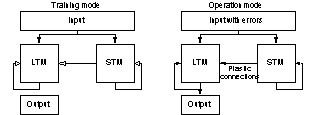
Figure 18 The system used in the
experiments of this section. Note the added plastic projection from the STM to
the LTM. The plastic projection is a full matrix (100x100) of weights.
To determine the
value of gSTM®LTM a trial and error process was used. Figure 19
shows 10 runs of the system, with different values of gSTM®LTM
in each run. G was set to the value 0.03 which corresponds to
approximately -3.5 on the
logarithmic scale of figure 19. If the value of g is set to a smaller
value, the retrieval-ratio of patterns 91-100 is decreased. If the g
is set to a larger value than 0.03 the retrieval-ratio of patterns 1-90 is
decreased. The dotted lines in figure 19 correspond to the performance of a LTM
and a STM.
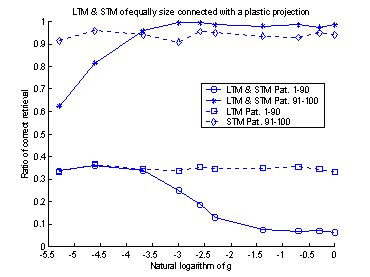
Figure 19 Ten runs of a system with plastic
projection. The systems retrieval-ratio is plotted against the logarithmic
value of gSTM®LTM. An
optimum can be found around -3.5. (The exponential of -3.5 is approximately
0.03.) Compare with figure 16.
Figure 20 shows the
retrieval-ratio of this system. The performance is very similar to that of the
system in section 4.1.2 were we had one-to-one connections. Comparing figure 20
with figure 17, one can se that the projectionSTM®LTM
interfere the LTM more then the one-to-one projectionSTM®LTM
did. If gSTM®LTM had been set to 1 this disruptive effect would
have been very prominent. The disruptive effect that the STM exerts on the LTM
depends on the number of elements that the projectionSTM®LTM
contain.
The use of a plastic projection
causes a small loss of retrieval-ratio performance, compared to the use of a
1-to-1 projection. This performance
loss is compensated by the versatility that the plastic projection provides.
Plastic projections allow
different representation, of the same data, in the systems different networks.
As I later will show, this can generate an increase of the systems performance.
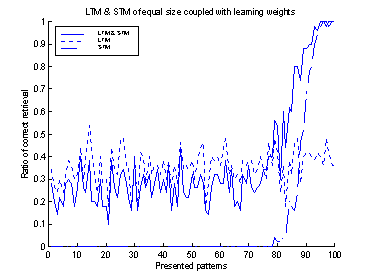
Figure 20 The performance for a system with
a plastic connection between a STM and a LTM of equal size. Compare with figure
17.
Two groups of neurons that are
far apart in the brain are usually very sparsely connected. This sounds
reasonable since it minimizes the hardware used. It can easily be understood
that all neurons in the brain cannot be connected to each other for volume
reasons. The experiment I preformed here was aimed at seeking out how the
performance is affected when connections between the LTM and STM are deleted.
In the experiment the
projection, that connected the STM to the LTM, was made sparse. The sparse
projection matrix was achieved through a random deletion of elements (deleted
elements were set to 1) in the projection matrix after the projection had been
trained. I made 4 runs of the system with different values of gSTM®LTM
in each run.
The influence of the STM on the
LTM was reduced when the number of connections was reduced. The influence was
then made stronger through an increase of gSTM®LTM.
The correlation between the sparseness of the projection matrix and the value
of gSTM®LTM
was of great interest. Figure 21 shows four plots with different values of gSTM®LTM.
In figure 21A a system with gSTM®LTM
= 0.03 is seen. The systems long-term memory storage capacity was not
compromised by the STM. About 40% of the elements in the projectionSTM®LTM
could be deleted before the system’s performance was affected. When finally all
elements of the projectionSTM®LTM had been
deleted, the systems performance was equal to the performance of the LTM alone.
In figure 21B, the value of gSTM®LTM
was increased to 0.1. The increase value of g made it possible to
eliminate 60% of all elements in the projectionSTM®LTM
without any major loss of performance. The decrease of the performance for the
last 10 patterns was steeper then in the previous plots. The plot to the lower
left, figure 21C, shows a simulation with gSTM®LTM
= 0.5.
The plot to the lower right,
figure 21D shows the performance of the system where gSTM®LTM
was set to 1. The STM suppress the LTM very effectively. Almost all elements in
the projection had to be removed before the suppressed retrieval-ratio of the
LTM could rise.
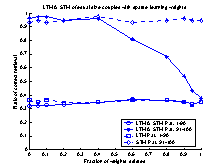
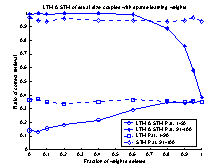
A B
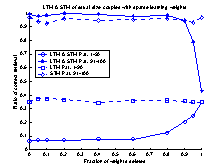
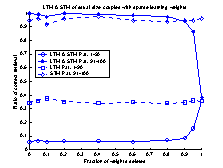
C D
Figure 21 The performance for 4 different
values of gSTM®LTM. The plot
in the upper left have g = 0.03,
the plot in the upper right have g = 0.1, the
plot in the lower left have g = 0.5 and
the plot in the lower right have g = 1. In
all of the four plots; in the left side of the plots all connections, between
the networks, are present, and to right side of the plots all connections,
between the networks, are removed.
It was very interesting to see
that more then 60% of the connection could be deleted without any major loss of
performance. This provides a hint that it would be possible to shrink the STM
without any loss of performance. The most interesting feature of the experiment
was that it clearly showed the need to have a scale-factor (gSTM®LTM)
between the connected networks. When gSTM®LTM
is set to 1 it is almost impossible to regulate the influence of the STM on the
LTM with the density of elements (connections) in the projection. This is seen
in figure 21D. The system, in figure 21D, act either as a STM or as a LTM.
An interesting question is what
happens if the patterns are represented differently in the LTM and the STM. If
the connections that provide the LTM with input are different to the ones that
provide the input to the STM. In the experiment I studied how the
transformation of the patterns affected the performance of the system. Was it
possible for the STM and LTM to cooperate with different representation of the
memories?
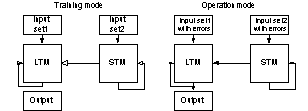
Figure 22 The system used two different sets
of patterns. In the experiments I studied how the STM could help the LTM to
retrieve the correct patterns, although it had been trained with a different
set of patterns.
In the experiment, I produced
one set of patterns that I used to train the LTM. I produced another set of
patterns that I used to train the STM. The hypercolumn structure of the input
patterns existed in both sets of patterns. The projectionSTM®LTM
was plastic. The constants of the system were set to the same values as in the
previous experiments.
In the experiment the
representation of the data differed in the STM and LTM. This meant that there
must occur a hetro-association between the patterns in the LTM and STM. This
was done by the plastic projectionSTM®LTM. It was
interesting to see that the performance of this system was almost better then
that of the system in 4.2.1, where we had the same representation of the
patterns in both of the memories. The slightly better performance can be
accredited to a more diverse and uncorrelated input.
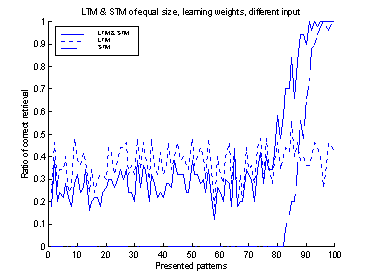
Figure 23 The performance for a system with
a plastic projection between the LTM and STM. Different representation of the
data was used in the LTM and STM.
A basic question was if two
separate networks with different plasticity works better then one single
network with two recurrent projections with different plasticity. It was
concluded that a separation of neurons into two networks with different
plasticity was good. It could also be
established that two networks with different plasticity could be made to work
together.
The concept with plastic
projection between the LTM and STM was seen to work. It was also established
that if a LTM and a STM were connected with plastic weights, the data could be
represented differently in the two networks.
The constant gSTM®LTM
was introduced to provide an instrument that could control the level of
influence between the networks of different plasticity. If gSTM®LTM
was set to 1, the STM had a dominant influence on the LTM. When gSTM®LTM
was set to 1 the LTM had problems to retrieve old patterns that were not stored
in the STM.
5.0 Properties
of connected networks
Chapter 4 was concerned with the
disruptive influence of the STM on the LTM. A paradigm of the system design in
chapter 4 was not to let the STM interfered the LTM capability to retrieve old
memories. The goal of the experiments in chapter 5 was to provide an
information base that could be used to design the systems of chapter 6, which
incorporated a STM that functioned as a working memory.
The LTM stores all memories,
although the most recent learned memories are not given precedence over older
memories. The role of the STM is to give the latest learned memories such precedence.
The STM can achieve this without having to store the latest memories. Remember
that the STM in chapter 4 stored whole patterns. Instead of storing the
patterns, the STM in 5.1 will hold pointers to the latest acquired memories.
Each of these pointers in the STM will be pointing at a particular memory in
the LTM. On retrieval of one of those particular memories the pointer will
become active and aid the retrieval of the memory.
In cognitive psychology chunking
is a popular concept. The compressed representation in the STM can be
considered as a chunk representing the content in the LTM.
The experiments in 5.1 were
designed to find out how a compressed STM could be constructed. Different
representations in the STM were tried. An investigation of different sizes of
the compressed STM was also performed. Note that when the system was in
operating mode, the activity was first propagated from the LTM to STM. Then the
activity was propagated back into the LTM. This was a big change from the
systems in chapter 4, where the STM was directly fed with activity.
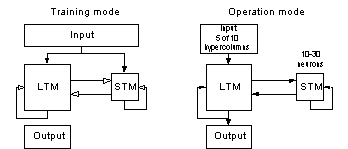
Figure 24 The design outline of the system
in 5.1.1-5.1.3. Note that during operation the activity in the LTM is
propagated through plastic projection to the STM, then the activity is
propagated back to the LTM. The STM consisted of 10-30 neurons.
The systems in 5.1 were
comprised of a LTM and a smaller STM. The LTM and STM were connected in both
directions with plastic projections.
Each of the systems was designed with three different sizes of the STM;
10, 20 and 30 neurons.
The plastic projections were
trained with aprojection
= 0.5. The constant g was set to 1 in both directions. The systems were trained
with 50 patterns. This implied that that the LTM were able to learn all of the
patterns.
The retrieval of the patterns
was initiated by presenting the system with 5 hypercolumns of the patterns. The
remaining 5 hypercolumns were left blank. (The activity for all units was set
to zero.)
Here, the STM was constructed
through a sub-sampling of the hypercolumns in the LTM. The STM with 10 neurons
was constructed simply through copying the content of the first hypercolumn in
the LTM. The STM with 20 neurons was constructed out of the first two
hypercolumns in the LTM. The STM with 30 neurons was constructed out of the
first three hypercolumns in the LTM. This meant that just a few of the
attributes (hypercolumns) of an object (memory) was accommodated by the STM.
These few attributes were stored with the full depth of detail retained.
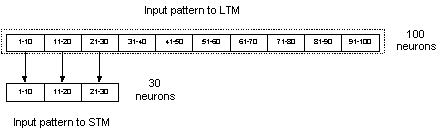
Figure 25 The outline for how the input
patterns were constructed. The input to the 30 neurons of the STM was
constructed out of the input to the first three hypercolumns of the LTM. When
the STM is constructed with 10 or 20 neurons 1 or 2 hypercolumns of the LTM are
used.
Figure 26 shows how the systems
retrieval-ratio with three different sizes of the STM. The system with only 10
neurons in the STM (dash-dotted line)
seemed to generate the best result.
A STM with only 10 neurons have
less influence on the LTM then a system with 30 neurons has. The STM consisting
of 10 neurons generate a projection on to the LTM containing 10x100 = 1000
elements, while a STM consisting of 30 neurons generate a projection with
30x100 = 3000 elements. A STM consisting of 30 neurons generates a more
distinct retrieval suggestion to the LTM than a STM consisting of 10 neurons. Setting the constant gSTM®LTM
to a value less then 1 can adjust the influence of the STM.
It was interesting to see that
even a small STM was able to help the LTM to activate the 10 latest patterns
correctly. This effect confirms the idea that the STM doesn’t need to contain
any information of the patterns, but instead can act as a pointer to the
patterns stored in the LTM.
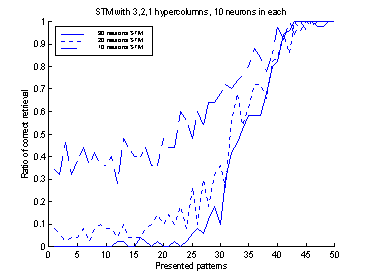
Figure 26 Three systems with different size
of the STM are shown in the plot. Presenting the systems with 5 out of 10
hypercolumns tested the retrieval-ratios of the systems.
The LTM was in this experiment
constructed through a compression of each hypercolumn in the LTM. The STM
contained the same number of hypercolumns as the LTM (10). Each hypercolumn in
the STM was comprised of 1, 2 or 3 neurons. This meant that all the attributes
of an object was stored in the STM, but with less detail. This approach was the
opposite of the approach taken in 5.1.1.
Note that the case where each of
the hypercolumns in the LTM was represented by a single neuron in the STM was
trivial. All of the neurons in the STM will always have the activity set to 1.
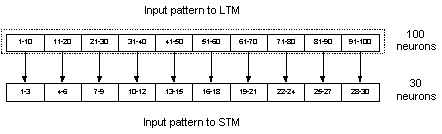
Figure 27 The outline for how the input
patterns were constructed. Data within each hypercolumn was compressed. All of
the 10 hypercolumns of the LTM are represented in the STM with 1, 2 or 3
neurons.
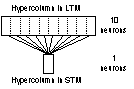
Figure 28 The figure shows how the hypercolumns
in the LTM was transformed to the hypercolumns of the STM. The left figure
corresponds to the case where the STM consisted of 30 neurons. The figure to
the right corresponds to a system with a STM of 10 neurons. Note that the right
figure is trivial.
The patterns stored in the STM
were highly correlated since each hypercolumn only had 1, 2 or 3 different
attribute values. Instinctively, this leads one to believe that the system
should have a poor performance; especially when the STM is composed of 10
neurons. A STM composed of 10 neurons divided into 10 hypercolumns is not able
to hold any information; all units would always be active. This was not the
case, as can be seen in figure 29. The retrieval-ratio of this system and the
system in section 5.1.1 was very similar. This can be explained with that much
of the information is stored in the plastic projection between STM and LTM. The
information stored in the STM seems to be of less importance.
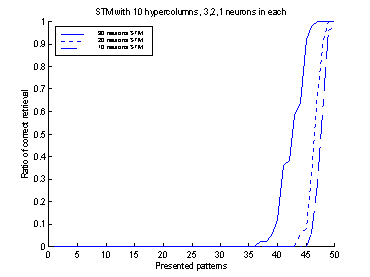
Figure 29 The performance for a system,
where each hypercolumn of the LTM was compressed from 10 neurons to 3 neurons
in the STM. Note that even the trivial case with 10 neurons, where all neurons
are active in the STM, can hold information.
Anders Lansner [personal
communication] have suggested that the relation between the number of
hypercolumns and the number neurons in a network, for maximal capacity, should
be
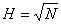
where H is the number of
hypercolumns and N the number of neurons. In 5.1.3 the compression of the LTM
was achieved through a compromise of sub-sampling and adopting a subset of the
hypercolumns. The number of hypercolumns in the STM was chosen to follow the
hypothesised relation.
The STM with 30 neurons had 6 hypercolumns.
The STM with 20 neurons had 4 hypercolumns and the STM with 10 neurons had 2
hypercolumns. Each hypercolumn in the STM was consisted of 5 neurons.
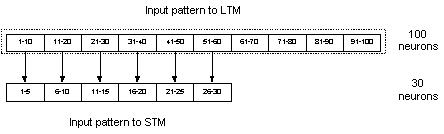
Figure 30 Shown here is the outline for how
the input patterns were constructed when the STM was made of 30 neurons. When
the STM was constructed with 20 neurons it had 4 hypercolumns, and when it was
constructed from 10 neurons it had 2 hypercolumns.
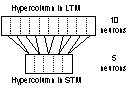
Figure 31 The hypercolumns of the LTM was
compressed to half its size in the STM. This applied for all STM, independently
of the number of neurons.
Figure 32 shows that the design
approach provides good performance. The STM constructed in this manner holds
more information then the STM of the two previous designs. It was interesting
to see that the size of the STM did not affect the performance of the system.
Comparing the results of the
experiments in 5.1, it is obvious that it is a good strategy to use a STM that
has a sparse representation.
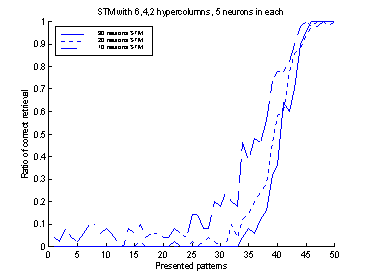
Figure 32 The performance for a system
where the STM is a subset of sub-sampled hypercolumns of the LTM. Note that the
size of the STM does not affect the performance of the system.
When faced with the task of
design there are often several parameters that can be adjusted in the system.
Section 5.2 contains an investigation on how some of the most important
parameters affect a system.
In these two experiments, the
focus was on how the plastic projection from the LTM to the STM affected the
performance of the whole system. As in 5.1 the activity of the LTM was
propagated to the STM and then back to the LTM. The phenomenon of interest in
these two experiments was the self-induced interference generated by the LTM.
In the two following experiments a plastic projection was used from the LTM to
the STM. One of the experiments used a plastic projection with a high
plasticity, and the other experiment used a plastic projection with low
plasticity.
The STM and LTM were of equal
size, 100 neurons. The STM had a
set to 0.5. The system had a one-to-one projection from the STM to the LTM. The
diagonal elements of the projection were set to the value 10. The choice of the
value 10 caused some impairment of the LTM ability to recall old patterns.
From the LTM to the STM there
was a plastic projection. In the case of low plasticity aprojection was set to 0.005 and in the case of high
plasticity aprojection
was set to 0.5. The plastic projectionLTM®STM
was scaled with gLTM®STM. Note that in this experiment, the gLTM®STM
constant applied to the projection from the LTM to the STM.
The system was trained with 50
patterns. The system was presented with noisy patterns to test the retrieval-ratio.
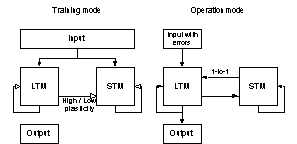
Figure 33 The system was used
to test the effects of the connection strength from the LTM to the STM. The
projection from the STM to the LTM was one-to-one and the diagonal elements
were set to the value 10. From the LTM to the STM there was a plastic
projection.
In figure 34 and figure 35 the
retrieval-ratio for the two systems is seen. The value of gLTM®STM
didn’t seem to affect the system as long as it was small. When the value of gLTM®STM
exceeded exp (3) »
20 a steep fall in performance for both the systems was seen. Most likely this
performance drop could be accredited to too much excitation of the STM. The
projectionSTM of the STM have up to this point, gLTM®STM
= exp (3) »
20, been able to suppress the activity imposed by the LTM.
The system with high plasticity
projectionLTM®STM had a constant performance for the last 10
patterns, independently of the value of gLTM®STM.
While the performance for the first 40 patterns slowly deteriorated as the
value of gLTM®STM increased. When gLTM®STM
exceeded 20 the performance of the system drastically dropped. The fact that
the performance for the last 10 patterns remained constant was logic since we
used a projection with high plasticity. As the influence of the LTM on the STM
increased, the memories in the STM were reinforced.
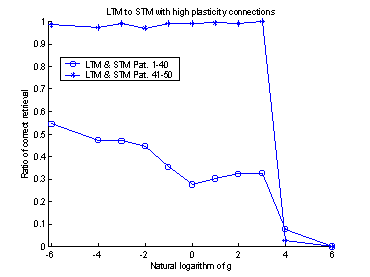
Figure 34 The system projection between
the LTM and STM had high plasticity.
The system with low plasticity
projections had a slowly deteriorating performance for the last 10 patterns as the
value of gLTM®STM increased. The performance for the first 40
patterns was independent of the value of gLTM®STM.
When gLTM®STM
exceeded 20, the performance drastically dropped as in the other system. The
fact that performance for the last 10 patterns slowly decreased, as the
influence of the LTM on the STM increased (larger gLTM®STM),
was logical since the system had low plasticity projection, which is not good
at storing the most recently learned patterns.
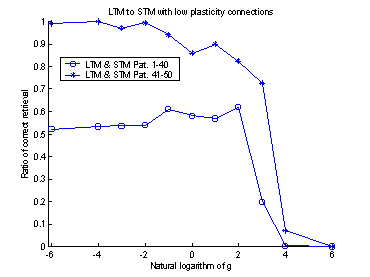
Figure 35 A system with a low plasticity projection
between the LTM and STM.
The projection in the direction
from the STM to the LTM is more important than the reciprocal projection, since
the state of the LTM is equal to the systems output. In this section I looked
at the influence of the projection from the STM to LTM was studied. First a
system with only one-to-one projection between the STM and LTM was studied.
Then a system with a LTM and STM of equal size with plastic projection was
studied. Finally a system with a compressed STM and plastic projection was
studied.
The first system was constructed
as the system in figure 15. The system had a fixed 1-to-1 projection from the
STM to the LTM, with the value 10. When the system was operated, noisy input
was fed directly to both the LTM and the STM. The LTM had a set to 0.005 and the STM
had a set
to 0.5. The systems were trained with 50 patterns.
Figure 36 shows the performance
of the system with a 1-to-1 projectionSTM®LTM.
When the value of the elements (weights) in the projection was smaller than exp (-2) » 0.1 a
decrease in the performance of the most recently learned patterns was seen.
This decrease seemed to be linear in respect to the logarithm of the weights.
It was also interesting to see that the retrieval-ratio of the older patterns
was not affected when the weights were smaller than 0.1.
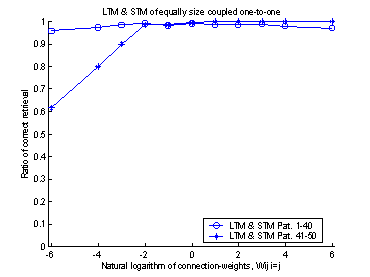
Figure 36 A system with a 1-to1 projection
from the STM to the LTM. The system was tested with different values of the
weights.
In a comparison between figure
36 and 37, it is seen that a 1-to-1 projection (figure 37) does not interfered
the LTM as much as plastic projection does.
The second system was
constructed with a STM and a LTM of equal size. The system design can be seen
in figure 18. The projectionSTM®LTM from the STM to
the LTM was plastic. The plastic projection was trained with aprojection
set to 0.5. The system was trained with 50 patterns. The retrieval-ratio was
tested with noisy patterns that were fed to both the LTM and the STM.
The performance of the second
system is shown in figure 37. The retrieval-ratio, for the last 10 patterns,
fall sharply when the value of gSTM®LTM
exceeds exp (3) »
20. The retrieval-ratio for the first 40 patterns starts to fall when the value
of gSTM®LTM
exceeds exp (-3) »
0.05.
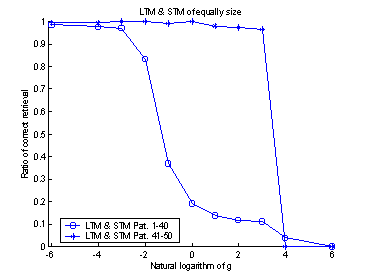
Figure 37 A system with equal size of the
LTM and STM. The plastic projection was trained with a set to the
value 0.5.
The last experiment was made
with a system similar to the one in figure 18. The difference was that this
system had a reduced size of the STM. When operated, the system was fed with
input directly to both the LTM and STM. The input to the LTM had errors, while
the input to the STM had no errors. The STM was made of 30 neurons. The input
to the STM was the same as the input to the first three hypercolumns of the
LTM.
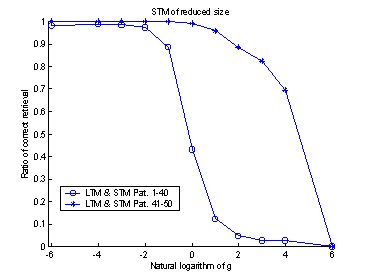
Figure 38 In this experiment the system
has a smaller STM then in the previous experiment.
The retrieval-ratio of the first
40 patterns is similar to that of the previous experiment. The retrieval-ratio
of the last 10 patterns starts to drop earlier than in the previous experiment.
The gentler drop of the retrieval-ratio of the last 10 patterns seen in this
experiment compared with the previous experiment can be accredited to the
reduced influence of the compressed STM.
How much information is needed
to retrieve a pattern in LTM and how does a STM affect the retrieval? These two
questions were answered by the experiment in 5.3. These two questions become
very relevant when one design a system where the patterns are dived into
individual modules, as in chapter 6.
The same system as in 5.1.1 was
used. The STM was composed of 30 neurons divided into 3 hypercolumns. The first
three hypercolumns of the patterns were stored in the STM. The system is shown
in figure 24. The system was run with four different values of gSTM®LTM.
The gSTM®LTM
scaled the projection from the STM to the LTM. The pattern retrieval was
initiated with 1 to 5 of the hypercolumns constituting the learned patterns.
In figure 39A one can clearly se
how the STM interfere the LTM. The STM have a positive effect on the retrieval
of the most recent patterns. Even though only the information of one
hypercolumn is presented to the system, it can retrieve the correct pattern.
In figure 39D the
retrieval-ratio of a system that does not have any STM is seen. If the system
is only presented with one hypercolumn, the retrieval-ratio becomes very low.
The system needs to be presented with 4 hypercolumns before the retrieval-ratio
becomes good.
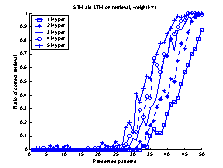
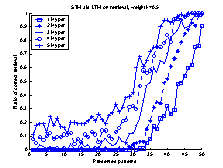
A B
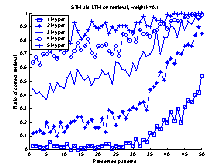
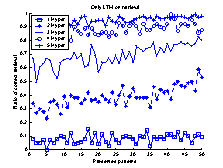
C D
Figure 39 Illustration of how the STM
affects retrieval of memories in LTM. Upper left plot have gSTM®LTM =1, upper
right gSTM®LTM =0.5,
lower left gSTM®LTM
=0.1.
In the lower right plot, the system has no STM.
This experiment was designed to
verify that a system composed of a LTM and STM put most significance on the
latest learned patterns. This means that if two patterns are very similar, and
the system is asked to retrieve one of these patterns it should retrieve the
most recently learned pattern. This also connects to the concept of STVB.
The system used was identical to
the system in 5.1.1 with a STM composed of 30 neurons. The patterns used as
input to the system consisted of two parts, called “Tag” and “Content”. The
input to the STM was the part of the pattern called “Tag”. The “Tag” can be
seen to represent a variable while the “Content” is representing the content of
the variable.
Figure 40 A pattern, with the parts tag and
content defined. The tag is represented by the first 3 hypercolumns, and the
content is represented by hypercolumns 4 to 10.
The system was trained with 50
patterns. The first pattern presented to the system, pattern A, was repeated a
number of times. The 48:th pattern was called pattern B. Patterns A and B were
very similar, their first six hypercolumns were identical.
When all of the 50 patterns had
been presented and learnt by the system, the first six hypercolumns of pattern
A (the first six hypercolumns of patterns A and B were identical) was presented
to the system. The system now had the choice of converging to pattern A or B.
The result is shown in figure 41. The system was run without the STM and the
result of this run is shown in figure 42. It may look strange that the
retrieval-ratio is sometimes larger than 1. The cause to this odd
characteristic is that sometimes the last 4 hypercolumns of the patterns A and
B are similar enough to cause both patterns, A and B, to be collapsed into one
single pattern.
The system in this experiment
performs a STVB task. The example of STVB given in 2.5.1, described how John
knew if he was talking to his grandfather, named Sven, or his friend, also
named Sven. To manage this task, John had to know which of these Sven’s he most
recently had met. The system in this experiment is presented with a similar
task. To refer to the example given in 2.5.1; the name of a person is in this
experiment is represented by the “Tag” and a physical person is represented by
the “Content”. Pattern A can be seen to represent John’s grandfather, named
Sven, and pattern B to represent John’s friend, also named Sven. The repeated
training of pattern A (representing the grandfather) can be seen as a long
conversation with the grandfather. The problem the system now face, is that
even if the systems has had a long talk to the grandfather, as soon as the
system starts to talk to the friend, the system knows directly that it is not
talking to the grandfather any more. To manage this task, the system needs to
swiftly change its references. The STM plays a crucial role in the swift change
of references.
In the first experiment where
the STM was enabled, the system almost only retrieved the latest learned
pattern, pattern B. (Figure 41)
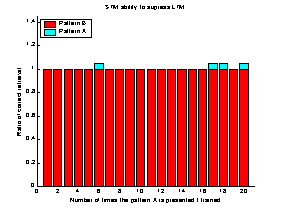
Figure 41 This histogram shows that the
system’s retrieval-ratio of the latest learned pattern, B. The retrieval-ratio
of pattern B was tested after different amounts of training with pattern A.
Pattern A and B had the same “Tag”. The figure shows, that if pattern A had
been trained 20 times, the system retrieved the most recent pattern, pattern B.
When the system’s STM was
disabled (Figure 42), the system only retrieved pattern B as long as the pattern A
had not been trained extensively. Once pattern A had been trained 3-4
times, the system almost never retrieved pattern B.
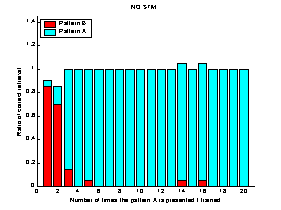
Figure 42 The performance the system, when
the STM was disabled. When pattern A has been repeated more than 2 times it is
almost impossible for the system to retrieve pattern B.
This experiment shows that a STM
can have a great impact on the system’s behaviour. White the STM, the system
could easily change the binding from the name variable to a new content.
Without the use of a STM, the system could not perform this task well.
The fundamental concept that the
STM does not need to contain the whole patterns that are stored in the LTM was
tried. 3 different approaches were taken to the design of the STM. It was seen
that the approaches that generated a sparse representation in the STM was
generally good. It was concluded that the STM only needed to be able to store
as many distinctive memory traces as the memory were supposed to hold. In our
case this meant that the STM should be able to hold about 10 distinct memory
traces. The information of particular patterns was stored in the projection
between the STM and LTM.
An investigation of how the LTM
and STM interfered each other was preformed. It was concluded that that the
interfering effect off the LTM on the STM was minimal. But if gSTM®LTM
was made bigger than 20 a drastic drop in the retrieval-ratio occurred.
It was also studied how the STM
interfered the LTM. If the two networks were connected with a 1-to-1
projection, the interference was minimal. The disruptive effect became much
larger when plastic weights were used. It was established that a smaller STM
interfered the LTM less then a large STM.
An experiment was performed to
investigate how much data a system of LTM and STM needed to retrieve a memory
(pattern). We concluded that a recent memory could be retrieved after
presenting the system with two hypercolumns. To retrieve an older memory the
system needed to be presented with information of four hypercolumns.
Finally we saw that a memory
system with both a LTM and STM, always gave precedence to the most recently
learned memories.
6.0 STM used
as working memory
Chapter 6 studied how a STM can
be implemented as a working memory. Two different systems were studied. The
system in 6.1 was based on a single STM and a single LTM. The system in 6.2 was
constructed with modules that were built of one LTM and one STM. These two
systems were tested on the task presented in figure 42.
There are 4 different places,
Place 1-4. In each place a box can be placed. There are 4 different boxes, Box
1-4. Each box has a certain content. If there are 4 boxes, there are 4
different contents, Content 1-4, one for each box. The task was to keep track of the boxes as they moved around to
different places. The working memory is supposed to hold the information that
Box 1 and Box 2 have switched place. The long-term memory holds the information
about what content each box has. It means that the systems need both a
long-term memory and a working memory to be able to perform the task.
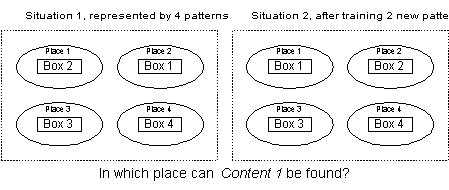
Figure 43 The system is first presented
with Situation 1, then Situation 2. After this two situation has been
presented, the system is asked to retrieve the place where “Content 1” is
stored. The 4 places are supposed to be well known. The 4 different boxes
and their individual content are also supposed to be well known. This
task is presented to the two systems in 6.1 and 6.2.
The purpose of this experiment was
to show that a STM could function as a working memory. The system was built
with one LTM and one STM. In section 6.2, a modified version of this system was
used as a module in a larger system.
The system used was almost
identical to the system in 5.1.1 with 30 neurons in the STM. There were two
differences. The first difference was that the input to the STM was taken from
hypercolumns 4-6 of the patterns. (Instead of the first 3 hypercolumns. This
difference is totally negligible.) The second difference was that only the
first 6 hypercolumns of the LTM were connected to the STM.
The patterns were composed of
three different parts as seen in figure 44. There were 4 different places.
There also were 4 different boxes. Each box had a particular content specific
to that particular box.
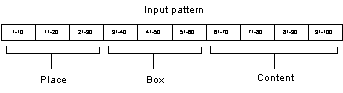
Figure 44 The input to the system had the structure
outlined here. Each Box had a certain Content. Each box
was placed in a certain Place. There were 4 places and 4 boxes with
their individual content.
The system was trained 2 times
with all possible combinations of boxes, and their content, in different places
(16 different combinations). The purpose of this training was to learn the
system each of the four “box-content” constellations. After these 2*16=32
patterns had been presented to the system, the system was presented with 10
patterns that contained noise. The last six patterns were more intricate.
Pattern 43-46 corresponds to situation 1 in figure 43. Pattern 47-48
corresponds to situation 2 (only the novelties in the new situation were learned)
in figure 43. All patterns, 1-48, are
documented in table 1.
|
Pattern No
|
Place
|
Box
|
Content
|
|
1-16
|
X
|
Y
|
Y
|
|
17-32
|
X
|
Y
|
Y
|
|
33-42
|
Noise
|
Noise
|
Noise
|
|
43
|
1
|
2
|
2
|
|
44
|
2
|
1
|
1
|
|
45
|
3
|
3
|
3
|
|
46
|
4
|
4
|
4
|
|
47
|
1
|
1
|
1
|
|
48
|
2
|
2
|
2
|
Table 1 This table shows the training
set of 48 patterns. Pattern 1-16 contain all possible combinations of X and Y
where X, Y Î {1,2,3,4}.
When the system was tested, it
was presented with the four different contents, Content 1-4. The system was
then asked to retrieve the corresponding “Box” and “Place” to each “Content”.
The system’s LTM associated each
“Content” to its corresponding “Box”. The “Box” in turn associated itself to
the correct “Place” using the working memory.
In table 2 the result of the run
is shown. Naturally, the retrieval-ratio of the content is 1. The
retrieval-ratios for the Box and the Place are also almost 1. Note that the
system has no problem too keep track of the last minute switch of Place between
Box 1 and Box 2.
|
Fraction of correct retrieval of:
|
Content
|
Box
|
Place
|
|
Box 1
|
1.00
|
0.98
|
0.98
|
|
Box 2
|
1.00
|
0.99
|
0.92
|
|
Box 3
|
1.00
|
1.00
|
0.99
|
|
Box 4
|
1.00
|
1.00
|
1.00
|
Table 2 The performance of the
system. The system was fed with a “content”, and then asked to retrieve
the place where this content was.
The same system was tested with
the STM disabled. The result is shown in table 3. The retrieval-ratio of the
content and box was still 1. This is what could be expected since the retrieval
of the “Box” is made with the LTM. It was interesting to see what happened to
retrieval of the “Place”.
The “Place” where Box 3 and Box
4 were put, was retrieved approximately 25% of the times. This corresponds to
the random frequency, when picking between four equal likely alternatives. The
same retrieval-ratio, 25%, of the “Place” were Box 1 and Box 2 were placed, was
expected. Instead I found the retrieval-ratio to be zero in these two cases.
This was strange.
Probably, the reason to the zero
retrieval-ratio was that the convergence was slower. All of the system in this thesis used a fixed convergence-time of
1 time-unit. The convergence-time in this case was probably up to 10
time-units. I did not depend the investigation into this matter, since I did
find relevant to the working memory.
|
Fraction of correct retrieval of:
|
Content
|
Box
|
Place
|
|
Box 1
|
1.00
|
1.00
|
0.00
|
|
Box 2
|
1.00
|
1.00
|
0.00
|
|
Box 3
|
1.00
|
1.00
|
0.26
|
|
Box 4
|
1.00
|
1.00
|
0.32
|
Table 3 The result of the system with
the STM disabled. Note that the system can not keep track of the switch between
box 1 and 2.
The aim of this experiment was
to show that a modular system could be designed and that this modular system
could perform equally well as the “integrated” system in 6.1. In this new
modular system, the representation of the Box/Content and the Place was
separated into separate modules. The approach, of constructing the systems with
LTM & STM modules, has many benefits over the systems with a single LTM
& STM. If the system is required to handle a new type of input / class of
attributes, it is easy just to add a module. And if the properties of a certain
class of attributes are altered, it is easy to alter the corresponding module.
The modular system is based on
two modules, each module contain a LTM and a STM. This system was tested on the
same task as the system in 6.1. Each of these modules is identical to the
system in 5.1.1. The STM contain 30 neurons and is connected to the LTM with a
plastic projection. Figure 45 shows the modular system. The bi-directional
projection between STM 1 and STM 2 has aprojection set to 0.5. The projections from STM 1 to LTM 2
and STM 2 to LTM 1 also have aprojection
set to 0.5. The bi-directional projection between LTM 1 and LTM 2 had aprojection
set to 0.005. The projections between LTM 1 and LTM 2 were made spares by
random deletion of 70% of the elements in the projections. The deletion of
these elements made the separation between the modules more clear. The aim was
to minimize the number of connection between the modules.
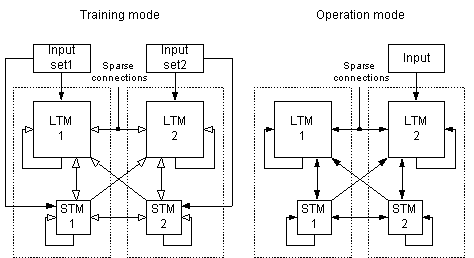
Figure 45 A system constructed of two
smaller systems from section 5.1.1. This system shows how bigger systems can be
constructed out of smaller modules. The system has two LTM connected with
sparse plastic projections, and two STM connected with plastic
projections.
In 6.1, all information was stored
in the single LTM. In this system, the “Place” and “Box / Content” memories are
stored in two separate LTM. Figure 46 shows how the “Place” memories are
represented in LTM 1. Figure 47 shows how the “Box / Content” memories are
represented in LTM 2. If the system is equipped with a third module, the
representation of the “Box / Content” can be separated.
Figure 46 The input to module 1 of the
system.

Figure 47 The input to module 2 of the system.
The system was trained with the
set of patterns described in table 1. The system was trained on each pattern
during 1 time-unit. On retrieval the system was fed with each of the four
”Content”. Retrieval (relaxation) was also made during 1 time-unit. The output
was taken from both LTM 2 and LTM 1.
The retrieval process of the
“Place” memory started with input of the “Content”. The system then used the
LTM 2
to activate the “Box” memory. The “Box” memory then activated the “Place”
memory, using the STM projections on LTM1.
The performance of the system
was good. Table 4 shows the result of
100 runs. This result shows that the modular design is working well.
|
Fraction of correct retrieval of:
|
Content
|
Box
|
Place
|
|
Content 1
|
1.00
|
1.00
|
0.99
|
|
Content 2
|
1.00
|
1.00
|
0.97
|
|
Content 3
|
1.00
|
1.00
|
0.99
|
|
Content 4
|
1.00
|
1.00
|
1.00
|
Table 4 The
performance of the modular system, when executing the task described in the
beginning of chapter 6.
Table 5 shows the result of a
run with the system where the two short-term memories, STM 1 and STM 2, have
been disabled. The retrieval-ratio of the “Place” memories is very poor. This
is expected, since the retrieval of these memories is dependent on the working
memory. If the result in table 5 is
compared with the result in table 3, one finds that the retrieval-ratio for
“Content 1” and “Content 2” is no longer zero, and the retrieval-ratio for
“Content 3” and “Content 4” has increased. As I earlier stated, these
differences can be traced back to the relaxation time and the different
networks structures and data representations.
|
Fraction of correct retrieval of:
|
Content
|
Box
|
Place
|
|
Content 1
|
1.00
|
1.00
|
0.14
|
|
Content 2
|
1.00
|
1.00
|
0.23
|
|
Content 3
|
1.00
|
1.00
|
0.40
|
|
Content 4
|
1.00
|
1.00
|
0.37
|
Table 5 The
performance of the modular system, when the STM 1 and STM 2 were disabled.
This experiment shows that
several Bayesian networks can be used in a modular designed system. The
experiment also, once again, proved the usefulness of a working memory. It
remains to be studied how these modular system scales.
It was shown that a STM, based
on a attractor network, could function as a working memory. We could also see
that a system with both a LTM and a STM could solve problems that would not
have been possible to solve with a system comprising of only a LTM or a STM.
Larger systems, based on modules
of LTM and STM, was constructed. We concluded that these systems could be
applied to solve the same problem as the smaller system, based on a single LTM
and STM. The advantages of the modular system were that its capabilities easily
could be extended and modified.
7.0 Discussion
The focus of this thesis was to
find out if a STM, based on fast changes in the synapses (weights), could be
constructed with the incremental, Bayesian learning rule. After it had been
established that it was possible to create a STM with fast changing synapses,
the focus turned on how the design of the STM could be refined. Several designs
of the STM were tried and evaluated. When this foundational work had been
completed, the attention was turned to the concept of working memory. The
question was, if it was possible to construct a system with a working memory
out of a STM and LTM.
Modelling the short-term storage
process with fast changing synapses proved to be successful. The idea that the
short-term memory process is similar to the long-term memory process allowed us
to adopt a concrete view of the STM. It also made it possible for us to
implement the STM as a high plasticity version of the LTM. If the short-term
memory process is based on some sort of persistent activity instead of fast
changes in the synapses, this model can still be applicable when modelling the
STM. The model proved that a STM could effectively be implemented, with an
existing LTM, with very few additional neurons.
On the network level, it was
established that Bayesian networks could successfully be operated with several
projections with different plasticity. This was a basic requirement to make it possible
for networks to cooperate. Useful insights on how to design a STM were
achieved.
Two different approaches were
tried. The first approach used a single population of neurons that had two
projections with different plasticity. The second approach used two networks
with different plasticity. This approach used 10% more neurons, but only 60% of
the connections compared to the first approach. The first approach could only
retain the last one or two memories while the second approach could retain about
10 of the most recently presented memories. In the cerebral cortex, both of
these types of memory may exist. The first type presented, with a short memory
span may exist in the visual regions of cortex. The second type of memory may
be found in the prefrontal regions of cortex where it may be used as working
memory [20].
The constant g
was introduced to control the influence a projection between populations
(networks) with different plasticity and size. The STM had larger influence on the
LTM, than the LTM had on the STM. The advantage of a network with low
plasticity was that more memories could be stored. The disadvantage was that
the memory became more sensitive to interferences. The size of the projections
also determined how much influence it had. A projection composed of many
weights had, naturally, more influence than a projection with few weights.
It was established that the
performance of a system with two projections, with different plasticity, was
improved if the system was divided into one high- and one low-plasticity
network. It was also established that the STM could be made much smaller than
the LTM. If the STM were to store the last 10 patterns, the STM needed to be
able to distinctively store a pointer or an address to each of these last 10
memories. If the STM had the same size of hyper columns as the LTM, the STM
needed 10 neurons to store 10 patterns. The number of neurons could be
increased even more if the hypercolumns in the STM were smaller than those of
the LTM.
Systems constructed with a LTM
and a STM were shown to be able to use their STM as a working memory.
This enabled these systems to perform operation that otherwise would have been
impossible. The working memory made it possible for the systems to perform “role
filling”.
Modules, or systems, of one LTM
and one STM were used to construct a large memory system. It was proved that
the functionality of a single module/system also was present in large system
composed of several modules. The system had more connections within each module
than between the modules. This characteristic, of localized connections,
conforms well to what have been seen in real neuronal systems [34].
There are several ways the
presented system can be interpreted, in a cortical sense. In the first
interpretation of the system a cortical hypercolumn correspond to a single
module. Each hypercolumn in the network correspond to a cortical column. The
individual neurons, of the system, correspond to a small number of inhibitory
and excitatory nerve cells. The other interpretation of the system is that a
module corresponds to a whole sensory area of the cortex, i.e. visual area of
cortex. Each hypercolumn in the network corresponds to a cortical hypercolumn.
And each neuron, in the system, corresponds to a cortical column.
An interesting concept to be
studied in the future is how the g factor affects the systems.
And also how a variable g factor can be used when a system is
extended with an attention control [35].
The incremental, Bayesian
learning rule, was created to deal with unlimited amounts of data. None of the
systems in this thesis were run with continuous streams of input and output
data. The systems were first put in training mode, and then they were put in
operation mode. To enable system to operate on continuous data streams, some
sort of regulating rhythm is needed that can control the switch between
learning and retrieval mode. Development of such an addition to the learning
rule is underway. The brain is thought to operate in the same manner, switching
between an input and output mode. The theta rhythm is thought to control the
switching between these two modes in the brain [36].
The working memory incorporates
a notion of time into the network. With the help of the working memory, the
system can keep track of the last occurring event. Even this small notion of
time proved to be useful, when the system were to perform tasks that were not
only those performed by associative memories. If the system is to become more
than just associative memory, it needs to be able to incorporate the dimension
of time.
8.0 References
1. Lynch,
G., 1999, Memory Consolidation and
Long-Term Potentiation, in The new
cognitive neurosciences. Bradford Books / MIT Press. p. 139.
2. Amit,
D.J. and N. Brunel, 1995, Learning
internal representation in an attractor neural network. Network. 6: p. 359.
3. Haberly,
L.B. and J.M. Bower, 1989, Olfactory
cortex: model circuit for study of associative memory. Trends Neurosci. 12(7): p. 258-64.
4. Hasselmo,
M.E., B.P. Anderson, and J.M. Bower, 1992, Cholinergic
modulation of cortical associative memory function. J. Neurophysiol. 67: p. 1230-1246.
5. Fransén,
E. and A. Lansner, 1995, Low spiking
rates in a population of mutually exciting pyramidal cells. Network:
Computation in Neural Systems. 6: p.
271-288.
6. Fransén,
E. and A. Lansner, 1998, A model of
cortical associative memory based on a horizontal network of connected columns.
Network: Computation in Neural Systems. 9:
p. 235-264.
7. Erickson,
C., B. Jagadeesh, and R. Desimone, 1999, Learning
and memory in the inferior temporal cortex of the Macaque, in The new cognitive neurosciences.
Bradford Books / MIT. p. 743.
8. Baddeley,
A., 1983, Working Memory. Philos.
Trans R. Soc. Lond. Biol. (302): p. 311-324.
9. Hopfield,
J.J., 1982, Neural networks and physical
systems with emergent collective computational abilities. Proc Natl Acad
Sci USA. 79(8): p. 2554-8.
10. Sandberg,
A., et al., 1999, An incremental Bayesian
learning rule,,, NADA, KTH.
11. Fuster,
J., M., 1995, Memory in the Cerebral
Cortex. London: The MIT Press.
12. Coltheart,
M., 1983, Iconic memory. Philos.
Trans R. Soc. Lond. Biol. (302): p. 283-294.
13. Tulving,
E., 1983, Elements of Episodic Memory.
Oxford: Clarendon Press.
14. Tulving,
E., 1987, Multiple memory systems and
consciousness. Hum.
Neurobiol. 6: p. 67-80.
15. Cohen,
N.J. and L.R. Squire, 1980, Preserved
learning and retention of pattern-analyzing skill in amnesia:
Dissociation of
knowing how and knowing that. Science. 210: p. 207-210.
16. Shepherd,
G.M. and C. Koch, 1990, The Synaptic
Organization of the Brain. New York: Oxford University
Press.
17. Cajál,
R.y., 1911, Histologie du Systéms Nerveux
de l'homme et des vertébrés.
18. Faggin,
F., 1991, VLSI Implementation of Neural
Networks, in An Introduction to
Neural and Electronic Networks.
19. Freeman,
W.J., 1975, Mass Action in the Nervous
System. New York: Academic Press.
20. Fuster,
J., M., 1989, The prefrontal cortex.
2 ed. New York: Raven Press.
21. Calvin,
W.H., 1995, Cortical Columns, Modules,
and Hebbian Cell Assemblies, in The
handbook of brain theory and neural networks. Bradford Books / MIT Press.
p. 269-272.
22. Churchland,
P.S. and T.J. Sejnowski, 1992, The
Computational Brain. Cambridge: MIT Press.
23. Eggermont,
J.J., 1990, The Correlative Brain: Theory
and Expirement in Neural Interaction.
24. Hebb,
D.O., 1949, The Organization of Behavior.
New York: John Wiley Inc.
25. Haykin,
S., 1999, Neural networks: a
comprehensive foundation. 2 ed: Prentice-Hall Inc.
26. Nadal,
J.P., et al., 1986, Networks of formal
neurons and memory palimpsests. Europhysics Letter. 1(10): p. 535-542.
27. Hertz,
J., A. Krogh, and R.G. Palmer, 1991, Introduction
to the Theory of Neural Computation: Addison-Wesely.
28. Lansner,
A. and Ö. Ekeberg. 1989. A One-Layered
Feedback Artificial Neural Network with a Bayesian Learning Rule. in Nordic Symposium on Neural Computing.
Hanasaari Culture Center, Espoo, Finland.
29. Lansner,
A. and A. Holst, 1996, A higher order
Bayesian neural network with spiking units. Int. J. Neural Systems. 7(2): p. 115-128.
30. Holst,
A., 1997, The Use of a Bayesian Neural
Network Model for Classification Tasks, in Dept. of Numerical Analysis and Computing Science, Kungl. Tekniska Högskolan, Stockholm.
31. Hubel,
D.H. and T.N. Wiesel, 1974, Uniformity of
monkey striate cortex: A parallel relationship between field size, scatter and
magnification factor. J. Comp. Neurol. 158:
p. 295-306.
32. Amit,
D.J., 1989, Modeling Brain Function: The
world of attractor neural networks. Cambridge University Press.
33. Dale,
H.H., 1935, Pharmacology and nerve
endings. Proc. R. Soc.
Med. 28: p. 319-332.
34. Johnston,
D. and S.M.-S. Wu, 1998, Fundamentals of
Cellular Neurophysiology: Bradford Books / MIT.
35. Hasselmo,
M., B. Wyble, and G. Wallstein, 1996, Encoding
and Retrieval of Episodic Memories: Role of Cholinergic and GABAergic
Modulation in the Hippocampus, in Hippocampus.
Wiley-Liss Inc. p. 693-708.
36. Kalat,
J.W., 1998, Biological Psychology. 6
ed: Brooks/Cole Publishing Company.
Figure
References
3. Unknown
URL, picture found on the Internet.
4. “Biological Psychology II:
Brain Structure and Function” http://psych.wisc.edu/faculty/pages/croberts/catonpic/topic4/cortex2.gif
5. Unknown
URL, picture found on the Internet.